第一讲 小麦遗传育种
第一节 小麦遗传育种概述
一、 小麦的分类与起源
(一) 小麦的分类
小麦属于禾本科(Poaceae),小麦族(Triticeae dumort),小麦属(Triticum L.)。小麦属包括一系列的多倍体物种:二倍体(2n=2x=14),四倍体(2n=4x=28),六倍体(2n=6x=42),现在被人类广泛种植的普通小麦(T.aestivum L.)为六倍体,染色体组为AABBDD,表1-1为小麦属的分类。
表1-1小麦属(Triticum L.)的分类
系 | 染色体组 | 类型 | 种 |
一粒系 Einkorn
| A | 野生 野生 带皮 | T.urartu Thum.乌拉尔图小麦 T boeoticum Boiss.野生一粒小麦 T.monococcum L.栽培一粒小麦 |
二粒系 Emmer | AB | 野生 带皮 带皮 带皮 裸粒 裸粒 裸粒 裸粒 裸粒 裸粒 | T dicoccoides Koem.野生二粒小麦 T dicoccum Schuebl.栽培二粒小麦 T.paleocolchicum Men.科尔希二粒小麦 T.ispahanicum Heslot.伊斯帕汗二粒小麦 T.carthlicum Nevski.波斯小麦 T.turgidum L.圆锥小麦 T.durum Desf.硬粒小麦 T.turanicum Jakubz.东方小麦 T.polonicum L. 波兰小麦 T.aethiopicum Jakubz. 埃塞俄比亚小麦 |
普通系 Dinkel | ABD | 带皮 带皮 带皮 裸粒 裸粒 裸粒 | T.spelta L. 斯卑尔脱小麦 T.macha Dek.et Men. 马卡小麦 T.vavilovi Jakubz. 瓦维洛夫小麦 T.compactum Host. 密穗小麦 T.sphaerococcum Perc.印度圆粒小麦 T.aestivum L. 普通小麦 |
提莫菲维系 Timopheevii | AG | 野生 带皮 | T.araraticum.Jakubz.阿拉拉特小麦 T.timopheevi Zhuk.提莫菲维小麦 |
茹科夫斯基系 Zhukovskyi | AAG | 带皮 | T.zhukovskyi Men.et Er. 茹科夫斯基小麦 |
(二)我国及新疆小麦的主要种类:
1.普通小麦,AABBDD,在全国品种数及面积分别占96%及98%,在新疆栽培最广泛。
2.园锥体小麦,AABB,品种数量占2%,在川、云、甘、陕、新等省区零星种植,60年代以后很少种。
3.硬粒小麦,AABB,品种数占1%,作通心粉,在新、蒙、陕春麦区曾种,塔额盆地有一定面积,硬粒率不达标。
4.密穗小麦,AABBDD,品种数占1%,穗椭圆,甘、云、晋、新曾种。
5.波兰小麦,AABB,品种极少,主要在新疆有(若羌古麦)
6.东方小麦(高拉山小麦),AABB,新疆有。
7.新疆小麦,AABBDD,又称稻麦子,新疆特有。
(三) 普通小麦的起源
二次异源多倍化过程:
(1)野生二倍体小麦T.urartu未知的或几个染色体组和现存山羊草属的Sitopsis组中某些种的染色体组相似的二倍体之间杂交形成双二倍体,即野生四倍体二粒小麦(T.dicoccoides)。
(2)野生四倍体二粒小麦(T.dicoccoides)与含有D组染色体组的二倍体方穗山羊草(Ae.Squarrosa)杂交形成异源六倍体小麦T.aestivum.
二倍体方穗山羊草 (Ae.Squarrosa),2n=2x=14,DD, D组染色体使六倍体小麦的面筋品质大为改善,并使穗轴易断特性消失。
六倍体小麦均为栽培种,没有野生六倍体小麦存在,所以六倍体普通小麦很可能是在栽培过程中产生的,而其易脱粒、圆粒特性则是后来突变的结果。
二、小麦的遗传特点
(一)普通小麦染色体组的多倍性
普通小麦染色体组为A、B、D3个染色体组,每个染色体组均为7条染色体,共计42条染色体(表1-2)。物种起源上的多源性使得普通小麦在染色体组上与小麦属和山羊草属的许多物种具有一定的亲缘关系,有一定的可交配性。
表1-2普通小麦的染色体组及部分同源群
部分同源群 | 染色体编号及染色体组 | ||
1 | 1A | 1B | 1D |
2 | 2A | 2B | 2D |
3 | 3A | 3B | 3D |
4 | 4A | 4B | 4D |
5 | 5A | 5B | 5D |
6 | 6A | 6B | 6D |
7 | 7A | 7B | 7D |
(二)普通小麦染色体的部分同源性
普通小麦染色体的部分同源群是小麦在进化过程中的一种剩余同源性,可能起源于共同的祖先。这种部分同源性的存在使得普通小麦在遗传性上趋于稳定和复杂化,首先是控制性状的基因增多,复等位基因广泛存在,性状变异幅度增大;其次是在每一个部分同源染色体群中可以出现染色体数量和染色体片断的增减,因此普通小麦通过染色体工程可以创造出缺体、单体、端体、三体、四体等染色体的数量变异和缺失、重复、倒位、易位等染色体的结构变异,这为进行小麦族内其他物种的基因向小麦中的转移以及进行小麦的遗传学研究提供了很大的便利。
(三)小麦的核外基因组
与其他植物种一样,小麦除细胞核内的基因组以外,在细胞质中的叶绿体和线粒体也含有相应的基因组。叶绿体DNA(cpDNA)和线粒体DNA(mtDNA)也对小麦的某些性状具有控制作用,例如小麦mtDNA对细胞质不育的控制作用等。同时由于核外基因组为母体遗传,所以有关小麦cpDNA和mtDNA图谱的研究对于阐明小麦及其亲缘物种之间的进化关系也具有十分重要的作用。
三、小麦的生态区域划分
(一)中国小麦的生态区划
见表1-3,共分为3个主区、10个亚区和29个副区。
1、北方冬麦区
长城以南、淮河、秦岭以北,眠山和大雪山以东,是我国小麦主产区。
(1)华北北部晚熟冬麦区:包括冀北、晋中、陕北、甘东、南疆小麦由这一区域引种成功。如唐山6898、新冬20、冀麦30(到新疆算早熟)
(2)黄淮平原中熟冬麦区:包括山东、晋南、冀南、苏北、皖北、关中、天水地区。
品种由关中向伊利引种容易成功。
2、南方半春性或春性麦区
淮河、秦岭以南、青藏高原以东。
(1)长江中下游平原中熟秋播关春性麦区:包括湘、鄂、赣、浙、苏中南、皖中南、陕西汉中。
品种引到新疆春播。如南大24129,阿勃(原产意大利)
(2)四川盆地早熟秋播半春性麦区。
(3)云贵高原早熟秋播半春性麦区。(云南昆明,元谋春麦南繁)
(4)华南山丘早熟秋播春麦区。
两广、闽、海南、(陆丰、三亚南繁。)
表1-3中国小麦生态区域的划分
主区 | 亚 区 | 副 区 |
冬(秋播) 麦区 | Ⅳ北部冬(秋播)麦区 | 10.燕太山麓平原区11.晋冀山地盆地区12.黄土高原沟壑区 |
V黄淮冬(秋播)麦区 | 13.黄淮平原区14.汾渭谷地区15.胶东丘陵区 | |
Ⅵ长江中下游冬(秋播)麦区 | 16.江淮平原区17.沿江滨湖区18.浙皖南部山地区19.湘赣丘陵区 | |
Ⅶ西南冬(秋播)麦区 | 20.云贵高原区21.四川盆地区22.陕南鄂山地西丘陵区 | |
Ⅷ华南冬(晚秋播)麦区 | 23.内陆山地丘陵区24.沿海平原区 | |
春(播) 麦区 | I东北春(播)麦区 | 1.北部高寒区2.东部湿润区3.西部干旱区 |
Ⅱ北部春(播)麦区 | 4.北部高原干旱区5.南部丘陵平原半干旱区 | |
Ⅲ西北春(播)麦区 | 6.银宁灌溉区7.陇西丘陵区8.河西走廊区9.荒漠干旱区 | |
冬、春兼播麦区 | Ⅸ新疆冬、春兼播麦区 | 25.北疆区 26.南疆区 |
X青藏春、冬兼播麦区 | 27.环湖盆地区28.青南藏北区29.川藏高原区 |
说明:1、划分三大麦区的两条界线:长城,淮河、秦岭。
2、划分生态区的重要依据:光、温、降水等生态条件;干旱、冻害、盐碱等自然灾害及病害;栽培制度、播种期及生育期。
3、生态区划分是相对的,冬春麦区只是根据播期分的,不表示其冬春性;同一生态区内条件及品种生态类型也十分复杂。
3、春麦区
长城以北、岷山、大雪山以西
(1)东北平原早熟春麦区
东三省、内蒙北部
(2)北部中熟春麦区
包括晋北、陕北、蒙中西、宁夏、甘中西、青海东部。
宁夏—北疆春麦区或兼种区引种。如宁春16,17
(3)新疆冬、春麦兼种区
(4)青藏高原春麦区
西藏、青海大部、川西、甘南
青海—新疆山间引种,如青春5号、高原465
(二)新疆小麦的生态区划
三大类型麦区:北疆、东疆春麦为主,南疆以冬麦为主,天山两侧冬春麦兼种,全疆冬春麦面积为2:1
全疆划为8个麦区:
1、冬麦区
(1)喀什—和田麦区
包括喀、和、克三地州,占全疆小麦面积的1/3以上,其中冬麦90%以上。
虽冬无积雪,但越冬条件好,中熟抗寒品种即可;两早配套,后期干热风,故强调早熟。
(2)阿克苏冬麦区
冬麦≥90% (棉花,水稻多) 有冻害,碱害
2、冬春麦兼种区
(3)伊力河谷冬春麦兼种区
伊力地区西—东,地势低—高,西部以冬麦为主,东部以春麦为主,冬麦占近60%
气候温和,春夏多雨、冬季多雪、水源充足,土地肥沃,锈病常发,
以当地自育冬麦为主(抗病),春麦有宁春16、17等
(4)乌苏—石河子—冒吉—奇台冬春麦兼种区
公路两侧积雪多,盆地边缘雪少,开春升温快,夏季高温、干热风;灌溉条件,机械化程度高。
冬麦奇台至乌伊公路沿线,西段多于东段,为棉花主产区,小麦面积日减。
春麦主要在奇台、吉木萨尔近山区,莫索湾或为与棉花轮作而种植春麦。
(5)轮台—库尔勒—若羌—且末冬春麦兼种区
巴州除焉耆盆地外各县、市冬春麦各半,轮台,若羌,且末种冬麦,库尔勒、尉犁因有冻害,故种春麦
3、春麦区
(6)阿勒泰——博乐——巴里坤春麦区,
包括阿勒泰地区、博州,塔额盆地、巴里坤、尹吾。
丘陵山区,气候温凉。500米以下无稳定雪层,800米以上积雪≥120天,不宜种冬麦,野燕麦,腥黑穗重(也有矮腥)
(7)吐鲁番—哈密盆地春麦区
吐鲁番地区、哈密市。
冬季无积雪,冬麦不便倒茬,只种春麦。
夏季炎热,有干热风;需早熟、抗干热风的品种。
(8)焉耆盆地春麦区
焉耆、和静、和硕、博湖四县。
无稳定积雪,盐碱重,不宜种冬麦;山间盆地冷凉,宜种春麦。
四、 全国及新疆小麦育种工作的发展
建国以来,小麦育种经历四个阶段:
(一)建国初期,50年代前中期:群众性良种评选。 如平原50,巴克扑克
(二)50年代中后期至70年代初期
全国以杂交育种为主,其他方法辅助,经几次大的品种更换。如碧蚂一号,9000万亩,因锈病生理小种改变淘汰―――教训:单一品种不行。
新疆以引进品种为主。如冬麦:新乌克兰83号;西北612;春麦:红星;阿勃;欧柔(喀什白皮――系统选育,当家十几年);墨麦(绿色革命标志之一)。
(三)70年代中期至90年代初期
全国株型育种(矮化)突破, 远缘杂交(小黑麦,小偃麦),花培、单倍体育种(京花1号,奎花1号)取得突破。
新疆以杂交育种选育出一批新品种,取代国外引进品种。如新冬2号、3号、14号、16号;新春2号、3号等。
(四)90年代中期以来
全国由产量育种转为品质育种为主,尤以河南、山东、黑龙江等省动作快而大,生物技术加紧探索在小麦育种上的应用。
新疆在超高产育种上进展大,如新春6号,新冬17号;早熟育种及品质育种也有突破,新冬22(奎冬5号)早熟,新冬18及新春8号优质。
第二节 小麦种质资源
小麦的种质资源包括栽培小麦、野生种和许多近缘植物。我国小麦种质资源的利用和开发在20世纪20年代就已经开始,然而,新中国建立之后才开始大规模有计划地展开。
一、种质资源的搜集与保存
1、国内种质资源的搜集与保存:
国内种质资源的搜集始于20年代,50年代初期大规模搜集, 80年代又进行一次补充搜集。
80年代还组织了3次大规模遗传资源考察,包括1979~1980年在云南地区,1981~1984年在西藏地区和1981~1983年在新疆地区的考察。
1986~1990年还专门对全国小麦野生近缘植物进行了考察,搜集小麦族遗传资源3, 000余份。到90年代,中国已搜集国内小麦属遗传资源19, 600余份,其中地方品种12, 879份,选育的品种(系)6700余份。
从各地搜集的品种看,我国栽培的主要是普通小麦,其次为圆锥小麦和密穗小麦,硬粒小麦在西北部有零星种植,而其他稀有种如东方小麦、波兰小麦都只有零散种植,此外不同地区还有一些特殊亚种,如云南小麦、新疆小麦和西藏半野生小麦。从种和亚种类型的丰富性看,有西多东少趋势,以新疆、西藏和云南为最多。
2、国外种质资源的搜集与保存:
20世纪20年代开始,以引进为主。
1921~1925年引进300多份,其中碧玉麦(亦称白玉皮)等曾在生产上应用;
1932年中央农业实验所引入潘氏(John Percival)世界小麦1700余份,从中选出南大2419(Mentana)、矮立多(Ardito)、中农28(villa Glori)在长江流域和黄淮南片应用;
1946年北京农业大学从美国引进2, 000余份,其中胜利麦和早洋麦在北方小面积种植并成为50年代华北地区骨干亲本。
40年代后期从美国引进的CI12203(甘肃96),也是50年代西北春麦区主要栽培品种之一。
到1990年底已从80多个国家和地区引进种质资源16, 500余份,除普通小麦外,也包括一些稀有种和野生近缘植物。
二、种质资源的评价和利用
金善宝、吴兆苏(1959)首先对国内小麦的种类、分布及特点进行了较全面的归纳和评价,认为我国小麦与世界各国比较,具有早熟、多花多实及某些特殊抗逆特性特点。
1、抗病种质资源的筛选和利用
有计划的鉴定始于50年代,尤以对条锈病抗性的鉴定最为系统和持久(国内条锈病大流行,碧蚂1号丧失抗条锈病能力)。中国农业科学院、北京农业大学等到1990年已发现和定名了29个条中号生理小种,90年代后期仍不断分化出新的类型;同时也不断筛选出新的抗源,这些抗源主要来自国外。
发现国内有些地方品种如平原50麦、蚂蚱麦等可能具有一定的水平抗性。初步明确含有pm2、pm4和pm6抗白粉病基因的材料在我国仍保持抗性,同时鉴定出一批新的抗源,如含pm21的材料。赤霉病的抗性鉴定虽然没有发现免疫品种,但也鉴定出抗性(主要抗扩展)强的材料,如江苏望水白、苏麦3号等。此外,对黄矮病、土传花叶病、吸浆虫等病虫害的抗性均进行过鉴定并筛选出一批抗(耐)性较好的种质资源。
2.优质种质资源的评价和利用
与国外品种相比,我国小麦品质情况:
(1)蛋白质含量不低:子粒蛋白质含量平均为12.57%(美国为12.97%),呈现北高南低;
(2)缺乏强筋力适于制作面包的硬质小麦和弱筋力适于饼干、蛋糕用的软质小麦,即“中间大,两头小”;
(3)普遍缺乏5+10等优质亚基,在GluAl、B1、D1位点上我国小麦以Null,7+8,2+12占大多数(85%以上),在D1位点普遍缺乏具5+10带(对面包品质有正向影响)的品种。但小偃6号(含14+15优质亚基)等优质小麦。
(4)我国还有相当数量资源,它们对改良面条品质具有相当潜力。
(5)我国还有一些特殊种质资源,如在糯蛋白位点上缺乏Wx-B1亚基品种和较为罕见的缺乏Wx-D1亚基的内乡白火麦等,与黑麦可交配性高的“中国春”,显性单基因控制的核不育基因ms2(太谷小麦),显性矮化基因的大拇指矮,显性矮秆基因Rhtl0的矮变1号(定位在4D染色体上)等。
在种质资源的利用上,除少数地方品种如平原50(水平抗病性)因其特殊种质在生产上直接利用外,大多数地方品种和外引品种是作为选择育种和杂交育种的亲本而间接利用。
外引品种如美国胜利麦和早洋麦;意大利品种(可能含中国早熟、多花亲缘的日本品种赤小麦血统而具有半矮秆、多花多实和抗条锈等特点,但因丰产性不强,不抗白粉病且品质较差而渐被取代);欧洲品种(l B/1 R小麦-黑麦代换系或易位系)洛夫林13、山前、高加索、牛朱特等在70~80年代成为我国育种中的骨干亲本;德国品种Heine Hvede在西藏也曾大面积种植(肥麦)并成为当地育种主要亲本,华北地区利用它育成的北京14、冀麦1号都在生产上发挥过重要作用;在亚洲种质资源中,引用最成功的是朝鲜的水源86(矮源),其次是印度育成的NP系列,巴基斯坦引进的墨巴系列也曾在局部地区利用。此外,拉丁美洲智利的欧柔(Orofen)也是我国小麦育种的一个重要亲本。
三、种质资源的创新
种质资源创新的手段主要有:
(1)不同类型品种间杂交;
(2)不同种属之间的杂交-远缘杂交;
(3)诱变育种。
品种间杂交育成的新的种质资源,我国做得比较突出的是矮源和抗源。
朝鲜的水源86→咸农39(咸阳农科所)→矮丰号系列(西北农学院);
有芒白2号×蚰包→半矮秆中间材料;
(意大利×印度亲源)×当地丰产品种→繁6(抗条锈和矮秆丰产,四川农业大学)衍生出 绵阳11、川育6号;
(矮丰3号×孟201)×牛朱特→矮孟牛(抗锈、矮秆、丰产,山东农业大学)衍生出 鲁麦1号
阿夫与台湾小麦杂交育成的抗赤霉病超亲的苏麦3号,已成为我国抗赤霉病育种的骨干亲本。
种间杂交主要以硬粒小麦与普通小麦杂交或再回交育成,例如中国农业科学院作物所优质小麦中8131—1就含有硬粒小麦血统。
第三节 小麦主要性状的遗传分析
一、小麦的形态特征:
多为质量性状,经济意义不大,但对育种和良繁有重要作用:
1.分类标志,(划)分变种(时),主要根据芒性,颖色,茸色,粒色
2.F1判别杂种真伪,如白穗×红穗--??
3.断别杂种后代的稳定一致性。
4.良繁中判别株行的典型性,一致性,鉴定品种纯度。
5.有的可对主要育种目标进行间接选择,如幼苗匍匐抗寒,红粒抗穗发芽等。
6.有的形态本身就是育种目标,如株高,茎粗、粒色、叶相。
(一)芒性:有利蒸腾和加速灌浆,防止风和鸟害,后期少量光合作用。
1.芒的发育:
分无芒、顶芒、有芒三类,无芒与顶芒为显性。
受四个显性抑制基因的控制:5A上的B1,6B上的B2,1D上的B3,4B上的Hd。
若B1B2B3Hd均纯合显性时,完全无芒,一对纯合显性为顶芒或曲芒(Hd),均纯合隐性为长芒。
2.芒色:
红、白、黑三色,红白与壳色一致。
有色(红、黑)为显性,有报导,1对或2对基因决定
(二)颖壳:
1.颖壳颜色:
有白、黄、棕、红、黑(普通小麦中多黑斑或黑条纹)
有色对无色(白、黄)为显性,一般F2呈单基因(3:1)或双基因(15:1)分离。
黑壳基因Bg位于1A上,红壳基因Rg位于1B上
2.颖壳茸毛:
绝大多数无毛,有利脱粒,扬场(如新春4号)。
有毛为显性,毛颖由位于1A上的一对显性基因Hg决定。
(三)籽粒
1.粒色
有白、红、紫、兰等色。
红皮对白皮为显性,受三对部分显性基因控制:3D上的R1、3A上的R2、3B上的R3。
种皮红色通透性差,休眠期长,抗穗发芽,皮厚些;
粒色的遗传呈部分显性并有累加效应,随显性基因的数目的增加而加深。
6R 5R 4R 3R 2R 1R 0R
最深红 暗红 深红 中红 浅红 淡红 白
紫种皮多从4倍体物种转移给6倍体小麦,呈显性,遮光使紫色不表现。
蓝色胚乳是由长穗偃麦草的基因决定的,如小偃兰粒,是普通小麦与长穗偃麦草杂交,染色体重组中,其中一条染色体被长穗偃麦草代换,称代换系,这条染色体上有决定蓝色胚乳的基因,它按部分显性遗传,有明显剂量效应。
2.籽粒硬度:
分为硬(角)、半硬(半角)、软(粉)
硬质的优点:制粉中可以得到大量制高级粉用的粗粉;制品流动性好;出粉率高;一般面筋好、色泽好(不绝对)。由1个或2个主基因控制,硬度遗传力≥90%,硬粒基因Ha位于5DS(短臂)上。
二倍体(AA)全是极软;四倍体(AABB)全是极硬;六倍体(AABBDD),从极硬――极软的一系列。
(四)茎杆
1.茎杆颜色:
可遗传的小麦紫杆,由位于7B上的Pc一个显性基因决定。黑麦的3R或4R与小麦置换或易位(染色体片断),也可引起小麦紫杆。
2.空心与实心
部分或全部节间空、实心,与抗倒伏有关,抗麦杆蝇。
一般硬粒小麦全部实心,园锥小麦穗下节实心,普通小麦多数空心,也有茎部或穗下穗节间实心或部分实心,茎杆实心可呈显性,隐性,受单一基因、互补基因或多基因控制。
(五)叶部性状
1.胚芽鞘:
芽鞘长度与小麦出苗早晚,顶土力有关,矮杆品种一般芽鞘短,不宜深播。
红(紫)、绿二色,红为显性(由位于7同源群上的三对基因RC1、RC2、RC3控制,随显性基因数增加,由绿到紫)
2.叶毛:
叶片(鞘)有毛对无毛呈显性,1对或2对基因控制,位于4A、5B上;
3.蜡质:
叶片及叶鞘的蜡质或粉露,有利抗旱,与抗蚜有关。
蜡质与2B、2D上的两位点有关,第一位点均可出现决定蜡质的基因。
(六)穗形:
六倍体小麦几个物种主要在三个基因位上存在差别:
5A上的Q 方形穗 穗轴坚韧
q 窄长形穗 穗轴易断
2D上的C 密穗
c 较稀长穗
3D上的S 籽粒长型
s 籽粒园型
二、高产性状
产量性状是个综合性状。
(一)产量结构
理论籽粒产量=单位面积穗数×每穗粒数×每粒重
高产品种结构三种类型(1000斤/亩)
多穗 型:(50-60万穗)×(25-30粒/穗)×(36-40克/千粒)
多穗为基础,力争提高千粒重,以保高产。
大穗型:(28-35万穗)×(40粒/穗)×(40-45克/千粒)
以增加第穗粒重为基础保证高产(1.3-1.6克/穗以上)
中间型:(40-45万穗)×(30-35粒/穗)×(40克/千粒)
以三因素同时并增为途径实现高产
在全国,南方麦区,多雨,少光不宜密植,穗分化时间长,宜大穗型;北方麦区干燥,多日照,穗分化及光照时间较短,宜多穗型。
新疆以中间型为宜,进一步提高应从多粒,大粒着手。(早熟的增穗数,中晚熟的增穗重)
产量性状多属微效基因控制的数量性状,一般不宜在早代选择,但也有差异。
1.每株穗数 遗传力很低。
2.穗粒数 与产量呈高度正相关(0.66),而千粒重与产量间仅0.25, 提高穗粒数是提高产量最重要而可靠的指标。穗粒数h2中等,40%左右,早代选择有一定效果;穗长与穗粒数呈正相关 ,而穗长h270%左右,可在早代选择。
3.千粒重 h2高( 70%左右),遗传主要受加性效应的基因控制。千粒重与每株粒数之间表示负相关,而又有补偿效应,故以穗粒重为产量育种的指标更为适宜。
(二)株高:
半矮杆品种推广是世界小麦产量提高的重要途径。
不是越矮越好,原因(?)(70-80cm为宜)
株高主要与节间数及节间长度有关,而节间数变化不大(5节),从抗倒考虑,应当基部节间短、粗、坚实、有韧性、弹力好。
株高遗传既受主基因的控制,又可能存在数目不等的修饰基因,多呈连续变异。 高×矮→F1多为中间偏高或超高亲,株高h2较高,50-70%,从F2起即可作为选择主要依据。
现已知矮杆的主效基因有二十多个,但得到广泛和最有效利用的是日本赤小麦(Akakomugi)的Rht8和Rht9和由达摩小麦(Daruma)为矮源育成的农林10号的Rht1 和Rht2。Ardito(矮立多)、 Mentana(南大2419)、 Funo(阿夫)、 Abbondanza(阿勃)半矮杆小麦→绿色革命。
农林10号×Brevor→Gaines(创世界高产记录)
(三)株型
是指地上部分的叶片、主茎、分孽和穗部在空间的存在状态,也就是植株受光的姿态。
现代概念已扩大到植株的一系列形态和生理性状的综合形式。
Donald(1968)曾提出理想株型:矮、独杆、少而小的直立叶,有芒大穗,繁茂种子根等,对此,争论很大,共同的理想株型及具体性状有一定片面性,同一个作物在不同地区,甚至在同一地区应有几个适当的理想株型。另外,应减少个体竞争,增大群体优势。
三、生理特性
(一)早熟性
1.意义: (1)增加复种指数;
(2)稳产(避免或减轻自然灾害)。
(3)可进行早晚品种搭配,缓和季节矛盾。
(4)减少与秋作物争水矛盾。
2.早熟的形态标志-抽穗期
抽穗期与成熟期呈高度相关,是完成阶段发育的可靠形态标志,且标准明确,记载简便,而且由于后期病害、干热风影响,成熟期往往不能真实反映正常情况下的成熟早晚。故育种上很少采用成熟期作为早熟性的指标。
3.早熟性的遗传:
早熟性受多基因控制,并广泛分布于每个染色体上。多以加性效应为主。
早熟性h2较高,达70%。早代有选择效果。
(二)抗旱性
干旱包括土壤干旱、大气干旱及混合干旱三类型。
小麦对干旱的抗性则有避旱(escape)、免旱(avoidance)和耐旱(tolerance)三类型。
1.抗旱品种的形态特征:
(1)植株繁茂,节间较长,茎较细,叶长而窄。
(2)分蘖较早,较多,分蘖生长快而集中,分蘖成穗率高。
(3)根系发达,根系分布广,吸收能力强。
2.抗旱鉴定的方法:
(1)直接鉴定:田间或盆栽观形态(干叶、卷叶、干芒),生长发育(生长速率,开花晚),灌浆饱满度
(2)间接鉴定:室内鉴定叶片含水量,叶片持水力,叶片水势
3.抗旱性选育的环境
单纯干旱协迫往往低产,应将F2放在中等条件下,后代分水、旱两圃或雨量不同的多点选育鉴定。
(三)抗寒性
低温对小麦的危害有:冻害(freezing injury)和冷害(chilling injury)。
小麦抗寒性是一种复杂的生理特性,与冬春性关系密切。一般情况下,强冬性及冬性品种比弱冬性及春性品种的抗寒性强,但二者不是完全一致,强冬性的品种不一定高度抗寒,如北欧品种。
抗寒品种形态指标:幼苗葡伏,叶片较窄,叶色较浓,冬前分蘖多,分蘖节深而大,叶鞘及分蘖节含糖量较高。
生理指标:新疆冬小麦要求分蘖节忍耐零下17-19度低温1-3天。
抗寒性遗传:多基因控制的数量性状,遗传率较高,早代选择有效。
(四)抗锈性
小麦锈病有条、叶、杆锈,
气传病害,周期大循环。
已鉴定出的条锈菌生理小种有条中1号至32号等生理小种;杆锈菌生理小种有17、19、21、21C1等12个;叶锈菌生理小种有1、2、3等三种类型。
抗锈性的遗传以垂直抗性研究的多,大多数是以主效基因控制的质量性状遗传。小麦抗条锈性的遗传率较高,早代应注意选择。
水平抗性为数量性状遗传,分离复杂。
目前已定名和描述的抗锈基因位点数目,抗条锈菌的有10个以上,抗杆锈菌的有30多个,抗叶锈菌的30多个。大多数已定名的抗锈基因都是小种转化性的(Johnson,1987)。
第四节 小麦育种途径和方法
一、育种途径和方法
(一)引种
从国内、外,70年代以前对农业生产发挥过很大作用。成功的:阿勃、阿夫、郑引1号、内乡5号、泰山1号等。
(二)系统选育
目前仍是小麦新品种选育的一个重要途径。成功的:东方红3号、博爱7203、杨麦3号等。
(三)杂交育种
是小麦育种中最为成功和成效最大的一项技术途径。
(四)远缘杂交
将小麦属的不同种及近缘种属中的抗病、抗旱、抗寒及优异品质的基因通过远缘杂交和染色体操作转移至小麦,丰富小麦育种资源,培育优良的小麦品种。
将黑麦的1R染色体片断导入小麦形成1B/1R易位系,对小麦的抗锈性改良发挥了巨大作用,如小偃6号,高产、优质、抗病。
(五)诱变育种
绝大多数用γ射线诱变育成,少数用X射线、热中子、快中子和激光诱变育成。成功的:山农辐63、鄂麦6号、鲁麦1号、新曙光1号、宁麦3号等。
在世界相关技术领域处领先地位。
(六)单倍体及其利用
原理:植物细胞的全能性。
主要手段:花药或花粉培养。
优点:减少杂种后代的分离,缩短育种年限。
我国是世界上第一个培育出小麦单倍体植株的国家。吴道芬的“京华1号”。
(七)杂种优势利用
目前小麦杂种优势利用的途径主要有三种:
三系法、两系法和化学杀雄法。
1. 三系法:细胞质不育或核质互作不育,如T型、K型、V型不育体系。
2. 两系法:
目前有两类方法:
(1) 只有不育系和父本系。靠标记基因区分育性,目前主要为蓝标不育体系;
(2) 光温敏不育两系法。
短日、低温―――雄性不育,用于制种;
长日、适温―――雄性可育,用于不育系繁殖。
3. 化学杀雄
70-80年代发现。
化学杀雄剂(Chemical hybridizing agent, CHA),又称化学杂交剂或杀配子剂(gametocide)。
目前只有三系法和化学杀雄法研制的杂交种进入了生产应用。
就育种途径而言,在2l世纪前半叶,常规育种仍将是小麦育种的主要技术途径,但同时要注意生物技术的应用,以扩大种质资源,提高育种效率。基因工程由于目的基因明确,且可跨越亲缘障碍,在一些基本技术环节完善后,在21世纪的后半叶将成为越来越重要的育种手段。
二、小麦育种的几个问题
(一)育种形式
1.多点育种(走马育种,适应性育种):一个单位在多个试点进行品种选育,或不同地区多个单位协作开展品种育种。提高材料利用率,扩大品种适应性。
意义和作用:不限于一个单位的多点试验,而在不同地区或不同国家之间的协作试验,共同利用杂种,不断交流,分别按目标选择。
2.穿梭育种:
例如 墨西哥:奥布雷贡 28。N 39m 下/11-5月
托卢卡 19。N 2640m 6-10月
中心材料送到巴西PH小于4土壤(溶铝)上鉴定耐铝性。巴西材料再回中心,往返穿梭。异地鉴定。
3.开放育种
中心是民间资助机构,各国在收获时可去选穗,美国一些大学或育种机关对公家和第三世界开放。
4.生态育种(克山农科所)
狭义生态育种:在某一生态条件下选育最近该条件的品种。
广义生态育种:在同一环境条件下,选育出适应不同自然区,不同生态条件的各类品种。
(二)杂种后代的处理:
1.选择程序宜灵活:系谱与混合法可以派生出许多方法,应因条件而定,人力,经费不足,搞混选,遇冻害、锈害就应单株选。
2.对杂种的纯度要求应适可而止,国内目前普遍对纯度要求很高。
(有限选择法,即F2、F3选择后就品系鉴定)
要有点剩余的异质性,不是机械的不纯,是遗传的不纯(较广的适应性和较持久的抗病性)
(三)育种规模宜大
美国俄勒冈大学3个点,每个点150-180亩几十万个系统“中心“每年5000个组合,选单株400万个;光小黑麦每年10000个组合,900亩。
第五节 小麦品质育种
一、小麦品质的概念
综合性概念。在各类谷物中,只有小麦含有独特的面筋,可以加工制成种类繁多的面食制品。
主要包括营养品质和加工品质两个方面,而加工品质又可分为磨粉品质(一次加工品质)和食品加工品质(二次加工品质)。在小麦收购、流通过程中,还经常采用籽粒形态品质的概念。
(一)营养品质(Nutrition Quality)
指籽粒中所含有的为人体所需要的各种营养成分,如蛋白质、氨基酸、糖类、脂肪、维生素、矿物质等。
小麦的营养品质好坏,不仅取决于小麦籽粒和面粉中各营养成分含量的多少,还取决于各营养成分是否全面和平衡。
(二)加工品质(Processing Quality)
将小麦籽粒磨制加工成面粉,再加工成各种面食制品,这个过程中对小麦品质的要求,称其为加工品质。
1.磨粉品质 (一次加工品质)
是指将小麦加工成面粉的过程中,加工机具和生产流程对小麦籽粒物理学特性(干粒重、容重、种皮厚度、硬度等)的要求。
普通小麦的磨粉品质要求出粉率高,粉色白,灰分少,粗粒多,磨粉简易,便于筛理,能耗低。要求小麦籽粒的容重高,籽粒大而整齐,饱满度好,皮薄,腹沟浅,胚乳质地较硬等。
2.食品加工品质(二次加工品质)
分为烘烤品质和蒸煮品质。
就烘烤品质而言,制作面包多选用蛋白质含量较高,面筋弹性好、筋力强,吸水率高的小麦及面粉;而烘烤饼干、糕点的小麦应选用软质小麦,要求面粉的蛋白质含量低,面筋弱,灰分少,粉色白,颗粒细腻,吸水率低,粘性较大。
对于蒸煮品质,制作面条的小麦一般为硬质或半硬质,要求面粉的延伸性好,筋力中等;蒸制馒头对面粉蛋白质含量和强度的要求比面包低,一般要求蛋白质含量中上,面筋含量稍高,中等强度,弹性和延伸性要好,发酵适中。
衡量小麦食品加工品质的标准主要取决于品种籽粒和面粉的最终用途(end use)。
二、小麦的品质分类和等级指标
商品小麦一般是根据皮色、粒质和播种季节进行分类的。
根据皮色,可将小麦分为红皮小麦和白皮小麦。红皮小麦的表皮为深红色或红褐色;白皮小麦的表皮为黄白色或乳白色;二者混在一起叫做混合小麦。
根据粒质可将小麦分为硬质小麦和软质小麦。硬质小麦的胚乳结构紧密,呈半透明状,亦称为角质或玻璃质;软质小麦的胚乳结构疏松,呈粉质状。
(一) 美国的小麦分类和等级
将商品小麦分为7类。
(1)硬质红粒冬麦(HRw) :面包小麦。蛋白质含量12%~14%,同时具有良好的磨粉和烘焙品质。该类小麦几乎占美国小麦生产量和出口量的40%。
(2)硬质红粒春麦(HRS) :面包小麦。蛋白质含量13%~16%,良好的碾磨和加工品质。出口量约占美国小麦出口总量的20%。
(3)软质红粒冬麦(SRw) :高产小麦。蛋白质含量10%左右,主要用于制作蛋糕、糕点、平式面包、饼干和一些小食品。
(4)硬粒小麦(Durum):四倍体小麦。硬度最高,用于制作通心粉和其它意大利式食品。
(5)软白麦(SW): 低蛋白质含量的小麦。蛋白质含量8%~10%,适合制作平式面包、糕点、馅饼、饼干和面条。
(6)不列级别小麦(unclassed wheat) : 指不能应用小麦标准中的规格进行划分的任何品种小麦。
(7)混合小麦(Mixed wheat): 由90%以下的一种小麦和10%以上的另一种小麦所组成,或由符合小麦定义的数种小麦混合而成。
美国农业部联邦检验署对美国七大类小麦分为五等,并对各项指标作了较为严格的规定。
表1-4 美国小麦的等级标准(徐兆飞等,2000)
等 级 | 磅/蒲式耳(最少重量) | 最大限度(%) | |||||||
硬红春麦白色密穗麦 | 其它品种 | 热伤粒 | 损伤粒 (总计) | 杂质 | 皱缩粒 破碎粒 | 疵项(总计) | 其它品种小麦 | ||
明显不同 | 其它品种 | ||||||||
一 | 58 | 60 | 0.2 | 2.0 | 0.5 | 3.0 | 3.0 | 1.0 | 3.0 |
二 | 57 | 58 | 0.2 | 4.0 | 1.0 | 5.0 | 5.0 | 2.0 | 5.0 |
三 | 55 | 56 | 0.5 | 7.0 | 2.0 | 8.0 | 8.0 | 3.0 | 10.0 |
四 | 53 | 54 | 1.0 | 10.0 | 3.0 | 12.0 | 12.0 | 10.0 | 10.0 |
五 | 50 | 51 | 3.0 | 15.0 | 5.0 | 20.0 | 20.0 | 10.0 | 10.0 |
注:1.损伤粒包括热伤粒;2.疵项包括损伤粒、杂质、皱缩粒和破碎粒,但总和不能超过疵项;3.任何非前三等的等级小麦可以包括不超过10%的其它品种小麦;4.其它品种包括明显不同的小麦。
(二)中国小麦的分类和质量标准
根据小麦的皮色、粒质和播种季节将小麦分为10类。
1.白色硬质冬小麦 种皮为白色或黄白色的麦粒不低于90%,角质率不低于70%的冬小麦。
2. 白色硬质春小麦 种皮为白色或黄白色的麦粒不低于90%,角质率不低于70%的春小麦.
3. 白色软质冬小麦 种皮为白色或黄白色的麦粒不低于90%,粉质率不低于70%的冬小麦.
4. 白色软质春小麦 种皮为白色或黄白色的麦粒不低于90%,粉质率不低于70%的春小麦.
5.红色硬质冬小麦冬小麦 种皮为深红色或红褐色的麦粒不低于90%,角质率不低于70%的冬小麦
6.红色硬质春小麦春小麦 种皮为深红色或红褐色的麦粒不低于90%,角质率不低于70%的春小麦
7.红色软质冬小麦冬小麦 种皮为深红色或红褐色的麦粒不低于90%,粉质率不低于70%的冬小麦
8.红色软质春小麦春小麦 种皮为深红色或红褐色的麦粒不低于90%,粉质率不低于70%的春小麦
9.混合小麦 不符合1至8各条规定的小麦。
10.其它类型小麦 分类方法另行规定。
以上各类小麦按容重分为五等(表2—3),低于五等的小麦为等外小麦。
表1-5 中国小麦质量标准(GBl351—1999)
等级 | 容重(克/升) | 不完善粒(%) | 杂质(%) | 水分(%) | 色泽、 气味 | |
总量 | 其中:矿物质 | |||||
1 | ≥790 | ≤6.0 | ≤1.0 | ≤0.5 | ≤12.5 | 正常 |
2 | ≥770 | ≤6.0 | ||||
3 | ≥750 | ≤6.0 | ||||
4 | ≥730 | ≤8.0 | ||||
5 | ≥710 | ≤10.0 | ||||
三、小麦品质的评价
小麦品质的好坏可从形态品质、营养品质、磨粉品质和食品加工品质等几方面进行评价。
(一) 形态品质(Shape Quality)
1.籽粒形状(Kernal Shape) :长圆形、卵圆形、椭圆形和短圆形等。以近圆形且腹沟浅的籽粒品质较优。
2.籽粒大小(Kernal Size): 以千粒重 “克”来表示。籽粒越大,越饱满,千粒重越高,且皮层比例较小,出粉率也较高。
3.籽粒整齐度(Kernal Evenness): 籽粒整齐度是指籽粒形状和大小的均匀一致性,可用一定大小筛孔的分级筛进行鉴定。籽粒整齐的品种,磨粉时去皮损失小,出粉率高,能耗也少。
4.籽粒饱满度(Kernal Plumpness) :分为饱满、较饱满、不饱满、秕瘦四级。饱满度好的籽粒磨粉时麸皮少,出粉率高。
5.籽粒颜色(Kernal Colour) :分为红色、琥珀色和白色。白粒品种在制粉时,出粉率较高,且面粉颜色也好,较受国内消费者欢迎,但易产生穗发芽。制粉业发达国家的绝大多数优质小麦品种都是红色。
6.籽粒硬度(Kernal Hardness) :分为硬质、半硬质和粉质三种。籽粒硬度与胚乳质地密切相关,对磨粉工序有较大影响。硬质小麦由于胚乳中淀粉与蛋白质紧密粘接,碾磨时耗能较多,但其胚乳易与麸皮分离,制粉时可得到较多形状较整齐的粗粒,流动性好,易于筛理,且出粉率高,面粉的麸星少,色泽较好,灰分含量较低;而粉质小麦则相反。
7.籽粒角质率(Percentage of Vitreous Kernal):角质胚乳和粉质胚乳。籽粒角质率是根据角质胚乳在小麦籽粒中所占的比例来确定的,与籽粒胚乳质地有关。角质率高的小麦籽粒硬度大,蛋白质含量高。中国把硬质小麦的硬质率规定为70%以上,把软质小麦的粉质率也定在70%以上。
(二)营养品质(Nutrition Quality)
1.蛋白质(Protein)
小麦籽粒的蛋白质分布很不均匀。胚占3.5%,胚乳占72.0%,糊粉层占15.0%,盾片占4.5%,果皮和种皮占4.0%。蛋白质含量变化很大。
小麦籽粒蛋白质根据其在不同溶剂中的溶解度,可将其分为清蛋白、球蛋白、醇溶蛋白和麦谷蛋白四种成分。清蛋白和球蛋白是可溶性蛋白,主要存在于胚和糊粉层中,其氨基酸含量较丰富,营养价值较高;醇溶蛋白和麦谷蛋白约占籽粒蛋白质总量的80%左右,并是组成面筋的主要成分,二者的含量及其比例,关系到小麦面粉的加工品质,也决定着小麦品质的优劣。
不同面食制品对蛋白质的数量与质量有不同的要求。
2.氨基酸(Amino Acid)
小麦籽粒蛋白质由20多种基本氨基酸组成,必需氨基酸和非必需氨基酸两类。其中,赖氨酸是第一限制性氨基酸,平均在0.36%左右,其含量只能满足人体需要的45%。因此,提高小麦蛋白质中赖氨酸含量是至关重要的。
3.淀粉(Starch) :小麦籽粒中最重要的碳水化合物。存在于胚乳中,约占小麦籽粒重的57%~67%,面粉重的67%,占胚乳重量的70%;;是面食制品的主要热量来源,并对籽粒和面粉的烘烤、蒸煮品质具有重要作用。
小麦淀粉粒在冷水中不溶解,用热水处理,可溶部分为直链淀粉,不溶部分为支链淀粉。直链淀粉含量的高低与馒头、面条的食用品质有关。直链淀粉含量适中或偏低的小麦制成的馒头体积大、韧弹性好、不黏,面条有韧性、不黏;而含量过高则制成的馒头体积小,韧性差,制成的面条易断。
小麦的营养品质还涉及纤维素(Celluloses)、游离糖(Free Succharide)含量、脂肪(Fats)、维生素(Vitamins)和 矿物质(Mineral Components)。
(三)磨粉品质(Milling Quality)
即一次加工品质,与小麦籽粒的许多性状有直接关系。磨粉品质好的小麦应为出粉率高,碾磨次数少,筛理容易,耗能低,粉色好,灰分含量低。
1.出粉率
单位重量籽粒所磨出的面粉与籽粒重量之比,即小麦粉重量占供磨籽粒重量的百分比。是衡量磨粉品质十分重要的指标。
高出粉率:籽粒圆大、整齐度好、种皮白薄、腹沟较浅、吸水率较高、籽粒偏硬等。
中国小麦的出粉率为79.0%~87.1%,平均为84.6%。一般来说,生产70粉时出粉率大于72%、生产85粉时出粉率大于86%的小麦品种受面粉厂欢迎。
2.灰分
是各种矿质元素、氧化物占籽粒或面粉的百分含量。是衡量面粉精度的重要指标。小麦皮层灰分含量为6%,而中心胚乳的灰分只有0.3%,因此,混入面粉的麸皮越多,面粉的灰分含量越高。
一般发达国家规定面粉的灰分含量在0.5%以下,中国富强粉的灰分含量为0.75%,标准粉为1.2%。新制定的小麦专用粉规定,面包用小麦灰分≤0.6%,面条和饺子粉≤0.55%等。
3.面粉白度
是磨粉品质的重要指标,已被列入国家小麦面粉标准的主要检测项目。
白度值与籽粒皮色、质地软硬、面粉粗细和含水量等有关。通常软质小麦的粉色比硬质小麦的粉色浅。面粉过粗、含水量过高都会使面粉白度下降。
新鲜面粉因含有胡萝卜素而常呈微黄色,贮藏时间长的面粉因胡萝卜素被氧化而使面粉变白。
中国小麦面粉(70粉)的白度为70%~84%。中国优质小麦规定一级大于76%,二级大于75%,三级大于72%。
(四)食品加工品质(processing quality)
不同类型食品对小麦籽粒和面粉品质的要求不同,各自都有相应的品质指标。
1.面筋(Giutenin)
小麦面粉经加水揉制成面团后,在水中揉洗,淀粉和麸皮微粒呈悬浮态分离出来,其水溶性和溶于稀酸的蛋白质等物质被洗去,剩留的有弹性和粘滞性的胶皮状物质称为面筋,用百分数表示。
小麦面粉之所以能加工成种类繁多的食品,就在于它具有特有的面筋。
面筋的主要成分是醇溶蛋白和麦谷蛋白。当面粉加水和成面团时,二者互相按一定规律结合,形成一种结实并具有弹性的像海绵一样的网络结构,即面筋骨架(glutenin net),其它成分,如脂肪、糖类、淀粉和水都包藏在面筋骨架的网络之中,使面筋具有膨胀性、延伸性和弹性等特性,从而可以制作面包、馒头、面条等各种面食制品。由此可见,醇溶蛋白和麦谷蛋白的含量高低及其二者的比例,共同决定着面筋的数量和质量,进而影响面粉的营养品质和加工品质。
小麦湿面筋含量一般在20%~35%之间。美国要求烤制优质面包所用的强力粉的面筋含量为36%~47%。中国新规定的强筋粉的湿面筋含量≥32%,弱筋粉的湿面筋含量≤22%。
2.沉降值(Sedimentation Value,SV)
将一定量面粉在量筒内与弱酸介质混合均匀,一定时间内蛋白质吸水膨胀所形成的悬浮沉淀数量的多少,以ml表示。是衡量面粉烘烤品质的一个重要指标。
大量研究表明,沉降值与食品加工品质呈显著或极显著的正相关,面粉的沉降值越大,表明面筋强度越大,烘烤品质就越好。
不同沉降值的面粉有不同用途,50毫升以上的面粉可以制作优质面包,20毫升以下的适于制作饼干,35毫升左右的适于制作馒头和面条。国外用于烘烤面包的小麦面粉的沉降值一般为60~80毫升。中国新规定的优质小麦标准中,强筋粉的沉降值应≥45毫升,弱筋粉应≤30毫升。
3.降落值(Falling Number)
降落值是指把装有一定量面粉悬浮液的粘度计管浸入热水器到粘度计搅拌降落入糊化的悬浮液中的总时间,以“秒”为单位表示。
反映了面粉中a-淀粉酶活性的大小,是检测小麦在收打和贮运过程中是否发芽的一项间接指标。降落值高的,表明a一淀粉酶活性低;反之则高。
根据小麦面粉降落值大小可将小麦分为三类:小于150秒的为发芽小麦,淀粉酶活性高,预示着面包心发粘;200~300秒之间为无发芽小麦,淀粉酶活性正常,可烤制出不干不粘有弹性的优质面包;大于300秒表明淀粉酶活性低,不利于酵母发酵,烤制的面包体积小,面包心干硬。
另外,粉质特性(由粉质仪(Farinograph)测得的面粉吸水率、面团形成时间、稳定时间、软化度、评价值等参数是评价面团流变学特性的重要指标)、 拉伸参数(用拉伸仪(Extensogl—aph)可测定面团延伸性和韧性的品质参数)、烘焙品质(面粉在制作面包、饼干及蛋糕等烘烤类食品过程中体现出来的,影响最终面制食品质量的品质性状-面包体积、比容、面包心的纹理结构、面包评分等,与特定的高分子量谷蛋白质亚基关系密切)与蒸煮品质也是重要的品质指标。
表1-6 国颁专用小麦品质部分指标(GB/T 17320-1998)
| 强筋 | 中筋 | 弱筋 |
蛋白质含量(%) | ≥14 | ≥ 13 | ≤13 |
湿面筋含量(%) | ≥32 | ≥28 | ≤28 |
沉淀值(ml) | ≥45 | 30-45 | ≤30 |
稳定时间(min) | ≥7 | 3-7 | ≤3 |
(五)不同面食制品对小麦品质的要求
表1-7 不同面食制品面粉的品质指标
项目 | 面包 | 方便面 | 馒头 | 面条 | 饺子 | 饼 干 | 蛋糕 | |
酥性 | 发酵 | |||||||
蛋白质含量(%) | >15 | >13.5 | 12~14 | >12 | >13 | <10 | — | — |
沉降值(毫升) | >45 | >35 | 25~30 | >30 | >30 | <18 | — | — |
湿面筋含量(%) | >35 | >30 | 25~30 | >28 | 28~32 | 22~26 | 24~30 | ≤22 |
面团吸水率(%) | >60 | >60 | 50~55 | — | >55 | 低 | 低 | 50~55 |
形成时间(分钟) | >6 | — | >3 | — | >4 | — | — | 1.0~1.5 |
稳定时间(分钟) | >10 | >6 | ≥3 | ≥4 | >5 | ≤2.5 | ≤3.5 | <1.5 |
降落值(秒) | 250~350 | >400 | ≥250 | ≥200 | ≥200 | ≥150 | 250~350 | ≥250 |
评价值 | >55 | — | 40~55 | — | >50 | — | — | — |
灰分(%) | ≤0.5 | <0.6 | ≤0.55 | ≤0.55 | ≤0.55 | ≤0.55 | ≤0.55 | ≤0.53 |
(引自高翔,200l)
第六节 2 1世纪小麦遗传育种工程技术展望
一、 21世纪我国小麦育种目标和途径
主体目标―高产、优质、高效。
1. 超高产
超级小麦概念的提出。认为在现有的品种类型上应进一步增大穗粒数和千粒重,降低单位面积穗数,向大穗类型发展。
表1-8是我国黄淮麦区未来超级小麦产量结构的几个优化模式。
表1-8 每666.7m2 650-700kg产量结构的优化模式
产量因素 | 每666.7m2穗数(万穗)×穗粒数(粒)×千粒重(g) | ||
优化模式 | 33.5×46.8×43.0 | 38.0×43.0×41.7 | 42.5×39.1×40.1 |
解的次数 | 115 | 165 | 99 |
频率(%) | 30.3 | 43.5 | 26.1 |
注意产量结构因子、群体冠层结构、生长发育节律、生物学产量以及同化物积累与运转特性等方面特异材料的创新。
2.优质
我国现有的绝大多数小麦品种的品质较差,面筋稳定时间较短,面筋强度较低是我国小麦品种品质较差的主要表现。
我国小麦品质状况目前是两头严重短缺,即适合于加工生产面包等食品所需要的强面筋小麦品种较少,适合于生产加工糕点类食品所需要的弱面筋小麦品种较少,这两类小麦目前主要靠国外进口。加强强面筋和弱面筋小麦品种的选育是我国小麦品质改良的主要目标。
同时,我国小麦消费的主要需求是面条和馒头等蒸煮食品,研究这些食品的品质标准,进行相应的专用品种的选育也是十分必要的。
3.高效
2l世纪的农业生产为高效质量型产业化生产,小麦品种不仅要具有较高的产量水平、优良的品质,还需要具有低消耗的高效特征。
21世纪小麦高效育种将主要集中在节氮、耐贫磷、抗旱等育种目标上。已有的研究表明,小麦不同品种间对氮素利用率为70~120kg/kg,品种间差异高达71.4%,氮素运转速率的品种间差异可达22%~78%,在同一土壤条件下,子粒含氮量的差异比其他器官的差异高48%~75%。在低磷条件下,磷高效的品种产量仅降低5%左右,而磷低效的品种产量可降低40%左右。在缺水的条件下,抗旱品种与不抗旱品种产量的差异幅度则更为显著。即使在水浇地,水分利用率(WUE)高的品种比水分利用率低的品种也具有较好的灌溉效率(IUE)。所以,进行小麦节氮、耐贫磷、抗旱、节水品种的选育具有十分巨大的潜力,这是小麦品种向高效型品种发展的一条重要途径。
二、现代生物技术与21世纪小麦遗传育种
现代生物技术可以在分子或细胞水平上对基因进行操作,从而定向改变生物的某些性状,同时打破物种间难以交配的天然屏障,使基因能够在不同物种之间相互交流、转移。21世纪生物技术必将对小麦生产起到非常重要的作用。
(一)细胞工程育种
小麦细胞工程主要包括花药培养、原生质体培养与体细胞杂交以及体细胞无性系变异等方面。
1.花药培养与加倍单倍体育种
花药培养是一种通过离体培养技术使小孢子脱分化,发育成愈伤组织或胚状体,进而再生出单倍体植株的技术。花药培养诱导的单倍体植株经过染色体加倍很快即可纯合成二倍体,从而大大缩短育种年限。
我国小麦花药培养技术取得了很大进展。具体表现在:
(1)研制出一批高效的新培养基:N6、马铃薯培养基、K3、c17、w14、麦合5号、癸等培养基。其中N6、马铃薯培养基、c17和w14培养基已经在全世界得到广泛应用,为小麦花药培养研究作出了很大贡献;
(2)对培养条件进行了改善与优化。提出了花药接种前低温预处理,接种后高温培养等有效的培养方法;
(3)广泛使用了桥梁亲本。通过以高绿苗诱导率的材料为桥梁亲本,使得杂种的花药绿苗诱导率得到明显提高;
(4)培养方式的改变。出现了固体培养、液体培养以及固体-液体培养等多种培养方式;
(5)提出培养基过滤灭菌,由甘露醇代替蔗糖等措施,使花药绿苗诱导率大幅度提高。
通过这些方面的研究改进,我国小麦平均花药愈伤组织诱导频率和绿苗分化率有了很大提高。80年代,我国小麦花药愈伤组织诱导率不足5%,绿苗诱导率1%左右,到90年代末,花药愈伤组织诱导率已经提高到10%以上,绿苗诱导率超过了5%。
染色体消失法诱导小麦单倍体的方法:利用小麦与亲缘较远物种如玉米、高粱、大麦、摩擦禾等杂交胚发育过程中外源染色体被排除掉的特性,通过杂种幼胚离体培养后分化出单倍体绿苗。
染色体消失法绿苗诱导率明显高于花药培养方法,而且在操作前可以对材料农艺性状进行选择,因而有着很大的应用价值。但目前这些方法还正处在研究与完善过程中,尚未得到广泛应用。
21世纪以花培为主的加倍单倍体育种的方向:
加大花药培养基础性研究力度。加大新高效培养基的研制、白苗形成的机理与影响因素研究以及农艺性状配合力好、花培诱导能力强亲本材料的筛选与利用等研究力度,使得花培绿苗诱导率有突破性的提高。远缘物种染色体消失法应完善操作程序,提高单倍体苗加倍频率,紧密结合育种,加快在育种中应用的步伐。
2.体细胞无性系变异与突变体筛选
体细胞无性系变异(somaclonal variation)是指由组织培养物再生植株所表现出的变异。在此基础上,通过在培养基中添加某种选择剂,就可以将组织或培养物中特定的细胞突变体筛选出来,用于遗传研究或育种。
小麦体细胞无性系变异现象相当普遍,变异所涉及到的性状也很广泛。这种变异具有诱导方便,稳定快,后代能基本保持供体的特性等特点,因而一开始就被广泛应用于小麦品种改良。利用体细胞无性系变异及突变体筛选技术已经筛选到了分别抗根腐病、抗赤霉病的突变体,抗盐突变体,高蛋白突变体等。
存在问题:体细胞无性系变异中有效变异很少,而且多为常见变异,因而利用价值有限。
(二)染色体工程育种
根据我国小麦染色体工程研究及应用现状和国情,以及世界染色体工程研究动态,在21世纪应当高度重视并加强染色体工程的研究与应用。应注意以下几方面的问题:
(1)加强染色体工程与传统育种技术、组织培养技术以及基因工程的结合;
(2)做好现有染色体工程材料的改造与利用研究:21世纪要对具有优良基因的材料进行改造、提高,重点创建小片段易位系,并进一步对现有易位系进行改良提高,使其更符合育种材料的要求,或培育新品种。
(3)围绕生产需求,突出重点:以小麦本身非常缺乏的基因如抗大麦黄矮病基因、抗赤霉病基因、优质基因等为主要目标,对小麦远缘材料进行大量筛选鉴定,并利用多种手段将这些基因快速转移到小麦中,为小麦育种和生产服务。
(4)加强染色体工程方面的基础研究:如改进、简化外源染色体鉴定技术,加大杀配子染色体、高配对基因等利用力度,做好基因定位研究等,降低染色体工程操作难度,提高准确性和有效性。
(三)基因工程
指人们按照预先设计的生物施工蓝图,把需要的目的基因经过体外切割、拼接与重新组合,然后引入受体细胞,并使其在受体内进行复制、表达,按要求改变受体细胞的遗传特性,然后由转化细胞再分化成具有预期新性状的工程植株。
1999年有7个转基因作物在12个国家种植,面积达到3 990×104hm2,我国1999年转基因作物种植面积为30×104hm2,成为世界上第四个转基因作物种植面积最大的国家。
小麦基因工程可分为3个部分:第一是目的基因的分离、纯化和体外重组;第二是遗传转化,包括将目的基因引入受体细胞并使其转化、转化体选择与植株再生等;第三是转化植株的鉴定与利用。
1.基因构建:小麦基因工程基因表达载体中经常使用的报告基因(reponer)有氯霉素乙酰转移酶基因(cat)以及B-糖苷酶基因(uidA)。最近,从水母中分离的绿色荧光蛋白基因(GFD)也被用作小麦遗传转化的报告基因。常用选择标记有新霉素磷酸转移酶基因(Npt一Ⅱ)(编码产物对某些氨基葡糖苷类抗生素如卡那霉素、新霉素等具有抗性)以及抗草甘膦的bar基因。常用的启动子(promoter)有花椰菜花叶病毒(CaMV)的35S启动子、增强caMV35S启动子、玉米来源的Ubiquitin基因(ubi)启动子、水稻来源的actin基因启动子。
2.遗传转化:可用来进行小麦遗传转化的方法有多种,但最为常用的有以下几种:
①基因枪转化法:将含目的基因的溶液与钨、金等金属颗粒混合,共同保温,使DNA吸附在金属颗粒表面,然后利用高压放电装置将金属颗粒加速,高速颗粒对细胞击孔,从而将目的基因带进受体细胞。基因枪法转化效率比较高。小麦首例转基因植株就是利用基因枪转化幼胚愈伤组织方法得到的。目前得到的小麦转化植株90%以上是利用基因枪直接转化幼胚的结果。
②电激法(electroporation):又称电激穿孔法。即利用高压放电产生的脉冲使受体细胞膜产生可逆的“微孔”,从而使外源DNA等大分子通过这些通道进入细胞,实现遗传转化的目的。小麦上也有少数几例利用电激法成功获得转化植株的报道。
③根癌农杆菌介导的遗传转化:该方法是发展最早、比较成熟的遗传转化方法。但农杆菌对小麦等大多数单子叶植物不感染或很难感染,一直无法用于小麦的遗传转化。但最近在这方面有了一定进展,Cheng等(1997)以幼胚为受体,利用根癌农杆菌介导获得了转基因小麦植株。但总的来说,这种转化方法转化效果非常差,还需要进行大量研究、探索。
④花粉管通道法(pollen—tube pathway):在授粉后一定时期内将外源总DNA或目的基因滴加或注入到子房,使其沿着花粉管生长时在花柱中开辟的通道进入胚囊,从而转化受精前后的卵细胞。该方法是由我国科学家自行设计的遗传转化方法,已经被广泛应用于棉花、水稻、小麦等作物,并获得了一批具有经济价值的稳定新品种(系)。利用这一方法已经得到一批抗白粉病、抗旱、抗大麦黄矮病等转化材料。
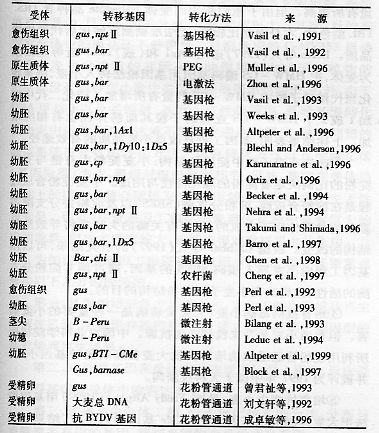
3.转基因植株的鉴定和利用:自1991年Vasil获得首例转基因小麦植株迄今,已经有20多例小麦转基因的实例(表1-9)。已经得到的转基因小麦包括抗病毒病、抗病虫、抗除草剂、改良品质等几个方面。
与其他重要作物相比,小麦基因工程研究及应用难度最大,进展最慢,仍存在不少问题与困难。首先,小麦基因数目庞大,调控体系很复杂,目的基因的分离、克隆难度很大。其次,目的基因在受体中的表达还不够理想。第三,缺少理想的载体系统与有效的转化方法。
表1-9已经得到的部分转基因小麦
(四)分子标记辅助选择育种
小麦遗传育种包括3个过程,即创造变异、选择所需基因型和鉴定与推广优良基因型。其中基因型选择是育种的重要环节。传统育种的后代选择主要以表现型来进行,方法简便、直观、经济,但易受环境条件、基因间互作(加性、上位性等)以及基因自身显隐性等多种因素干扰,不得不进行多年多点试验或在特殊环境条件下进行选择,预见性较差,工作量大,周期长,妨碍了育种进程。
通过与目标性状紧密连锁的遗传标记可以对目标性状进行跟踪选择,提高传统育种选择水平。
遗传标记辅助选择必须具备三方面的要求:
① 标记必须与目标性状共分离或紧密连锁;
② 能够对大群体进行筛选、评价;
③ 技术要可靠,重复性好,经济,对人类无害。
1.遗传标记的类型
目前遗传标记有三大类,即形态标记、生化标记和分子标记。
2.分子标记的优越性:
① 直接以DNA的形式表现,在植物各组织、器官,各发育时期均可检测,完全不受季节、环境及材料的限制;
② 分子标记数目极大,遍及整个基因组。从理论上讲,可以找到与任何目标性状紧密连锁的标记;
③ 多态性高。植物自然存在许多等位变异,不需要专门创造特殊的遗传材料;
④ 分子标记不影响目标性状的表达,与不良性状无必然的连锁关系;
⑤ 有许多分子标记呈现简单孟德尔遗传,表现为共显性,表型与基因型统一,能够有效识别纯合基因型与杂合基因型;
⑥ 分子标记间无干扰,可以在同一试验内同时利用很多种分子标记对很多个目标性状进行选择。
分子标记技术发展十分迅速,21世纪必将给小麦育种带来革命性的变化。
3.分子标记的种类
①以DNA分子杂交技术为基础的RFLP标记;
②以DNA体外扩增技术(PCR)为基础的分子标记,如RAPD、SSR、AFLP等。
表1-12已经被标记的小麦基因
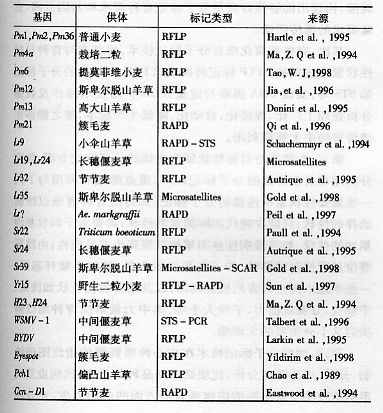
分子标记在小麦遗传育种中的使用:主要用于绘制指纹图谱,检测种子质量,保护知识产权;对骨干亲本进行分类,确定亲本间的遗传关系,科学配置杂交组合;种质资源创新与鉴定以及杂交后代目标性状的鉴定筛选(分子标记辅助育种,marker—assisted—breeding)等方面。
在21世纪,小麦分子标记技术及育种应用前景十分广阔,但必须切实抓好以下几个方面的工作:
(1)继续做好小麦遗传图谱的绘制工作,研制并采用新的分子标记技术,填补遗传图谱中的大缝隙,构建出能够很好满足分子标记育种需求和基因分离、克隆要求的遗传图谱。
(2)完善并简化现有分子标记技术,将现有与育种目标性状紧密连锁的RFLP标记转换成以PCR为基础的分子标记如STS等;实现DNA提取与定量,PCR扩增技术操作及数据分析处理工厂化、规模化、自动化,大规模利用。
(3)加强分子标记技术在核心种质资源的指纹图谱绘制、分类、遗传距离分析,优质高产新品种的选育,抗病虫及病毒病新种质与新品种的快速选育等方面的应用力度,充分利用有限的财力、物力,把重点放在对小麦生产影响较大,而且又能够通过分子标记技术解决的问题上。
(4)分子标记与传统育种紧密结合,充分利用传统育种者丰富的育种经验,最大限度发挥分子标记准确高效的特性,快速选育优良小麦新品种,为我国小麦生产作出贡献。