第七章 细菌和噬菌体的重组和连锁
计划学时6hr
原核生物 prokaryotes
无明显的细胞核,不进行RD!
病毒:
无细胞结构,基因传递方式是非RD!式
第一节 细菌和病毒在遗传学研究中的地位
一、关于细菌和病毒
1.细菌
● 单细胞的原核微生物
● 形状(球形、杆形、螺旋形)、大小变化很多
● 遗传物质是一个大型的环状核酸分子——基因带裸露的DNA分子 genophore
● 繁殖方式:无性的裂殖法,有性生殖——菌落 clone
营养缺陷型 auxofroph
由于基因的突变,细菌的菌落或菌株(strain)丧失产生某种营养物质的能力(通常是AA)称为营养缺陷型
原养型(proto froph)
相对于缺陷型的生产出型菌株,称为原养型
2.病毒
● 无细胞结构
● 无代谢功能——寄生在活细胞内
● 蛋白质外壳 + 核酸构成 bacteriophage
● 动物病毒,植物病毒,细菌病毒(噬菌体——phage)
二、细菌和病毒是遗传学研究的好材料
结构简单,繁殖力强,世代时间短
培养要求和生理状态的变化多
大多能在一定的人工培养条件下生长繁殖
①细菌大都能合成全部的AA和维生素
②基因作用的研究,营养缺陷型——MM +?AA V 生长
③突变频率很低,但选择容易,如抗链霉素突变
④研究基因的精细结构(phage)
某一基因内的大量不同位点(sites)的突变型可以通过重组分析测定它们的确切位置
⑤代谢调控问题的研究应用到高等动、植物中,推动其问题的研究
⑥便于建立纯系,长期保芷
几个符号 + 野生物
- 缺陷型或突变型
s 敏感 sensitive
r 抗性 resistant
第二节 细菌的遗传分析
大肠杆菌是用的最为广泛的遗传学实验材料,人工培养极方便(简单的培养基上生长,含有盐类+葡萄糖)
一、细菌杂交实验 minimal medium(MM 基本培养基)
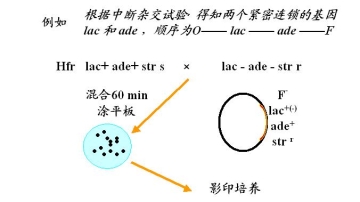
1.杂交亲本介绍 大肠杆菌K12中的两个菌株A和菌株B
菌株A: met— bio— thr+ leu+ thi+
甲硫AA 生物素 苏AA 亮AA VB1(硫胺素)
培养菌株A MM + met + bio
菌株B: met+ bio+ thr— leu— thi—
培养菌株B MM + thr + leu + thi
2.杂交实验一
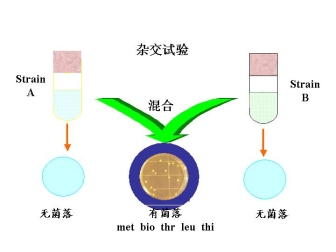
引出的问题:
能在基本培养基上生长的这种细菌
?①是由于营养上的互补—— 一种物质从一个菌株的细胞中泄露漏出来,而为另一菌株的细胞所吸引?
?②菌株A和菌株B发生杂交,交换遗传信息(genetic information)
出现重组,产生met+ 、bio+ 、thr+ 、leu+ 、thi+的细菌
原因可能有二
其一:某菌株的遗传物质进入另一菌株的细胞内,发生重组——转化(无细胞接触)
其二:两菌株的直接接触,传递遗传物质,发生重组——接合
解决这些问题的办法:设计U管试管
3.U型管试验
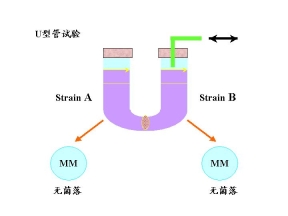
实验说明:在基本培养基上出现能够生长的菌落
①一定有两个菌株的直接接触
②一定遗传物质的重新组合
二、F因子
(两个菌株如何接触?又如何发生遗传物质的重新组合)
1.Hayes的杂交试验
菌株A:met— thr+ leu+ thi+
菌株B:met+ thr— leu— thi—
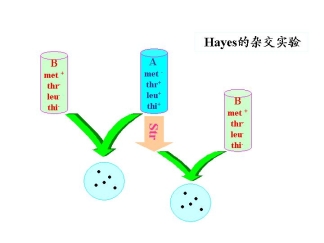
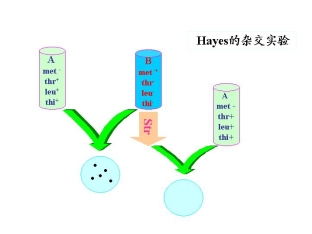
实验说明:
①大肠杆菌中遗传物质的交换不是交互的
②菌株B作为遗传物质的受体(recipient)
菌株A作为遗传物质的供体(donor)
供体经str处理后不能分裂,但能转移遗传物质,受体可以接受遗传物质,因为无str处理,可以分裂因而产生菌落。
2.F因子
——细菌细胞内的一种微小的可转移的因子,又称性因子或致育因子(sex or fertility factor)
⑴F因子的特性
● 环状的DNA分子,其上有能形成性伞毛的基因
 ● 独立增殖(复制)
● 独立增殖(复制)
● “核”DNA以外的DNA
● 游离于核外,或整合在“核”DNA内
● 具有转移的能力
● 供体细胞中具有
⑵大肠杆菌的性别
F+ 具有F因子,作为供体,菌体表面有性伞毛
F— 无F因子,作为受体,菌体表面无性伞毛
⑶F因子的转移
F+ × F+ → 无接合 因为F因子改变细胞表面的构造,防止等接合
F— × F— → 无接合 因为无F因子,无性伞毛
F+ × F— → F+ + F+ 因为F因子可以复制并从供体传递给受体
F+ → F— 特殊处理(如吖黄素),F因子丢失
三、高频重组与中断杂交技术
1.高频重组(high frequency of recombination)
⑴Hfr菌株
为何称Hfr菌株为高频重组菌株
 F+ × F— → F+ + F+ (重组子频率很低)
F+ × F— → F+ + F+ (重组子频率很低)
Hfr × F— → Hgt + F— (但重组子频率很高)
⑵Hfr的特点
①有F因子,但整合在“核”基因组中
②Hfr与F—较F—很少转变为F+细菌,但重组频率很高
2.中断杂交技术
(1)中断杂交实验
菌株A(供体)
Hfr | thr+ | leu+ | azir | tonr | lac+ | gal+ | strs |
苏AA | 亮AA | NaN3 | NaN3 | 噬T1 phageT1 | 乳糖 | 半乳糖 | 链霉素 |
菌珠B(受体)
F— thr— leu— azis tons lac— gal— strr
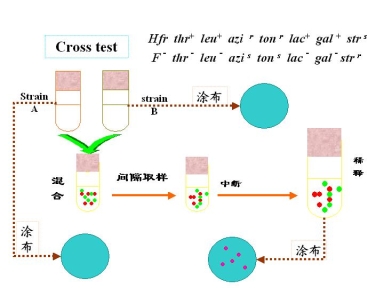

如果有菌群生长表明Hfr上的非选择性标记基因进入F—中
在某选择培养基上出现的菌落百分数愈高,说明该基因出现的时间愈早:根据时间的长短和出现供体基因的顺序作图——连锁图
(2)连锁图和中断杂交技术
根据供体基因进入受体细胞的顺序和时间绘制的遗传图称为连锁图,绘制连锁图的技术称为中断杂交技术
实验说明的问题
①Hfr基在按一定的顺序依次进入F—细胞原点(origin)是Hfr转移入F—的起点
②Hfr×F—杂交二小时后
![]()
Hfr+Hfr说明致育因子(F因子)最后进入受体
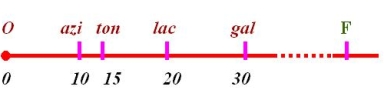
四、大肠杆菌的环状染色体
表7-3几个Hfr菌株的基因顺序
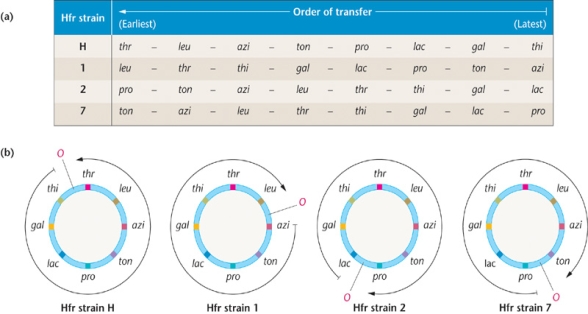
看出:基因转移顺序转移起点转移方向各不相同
![]()
![]() 但有规律可寻O F
但有规律可寻O F
比较四种Hfr菌株后如果不考虑起点和F因子的位置,可以很容易发现
①大肠杆菌的“染色体”是环形的
②在不同的Hfr品中只有O和F的位置不同
原因何在?即F因子插入的位置和插入方式
五、F因子整合到细菌染色体的过程
——解决一个细菌染色体 + F因子——整合
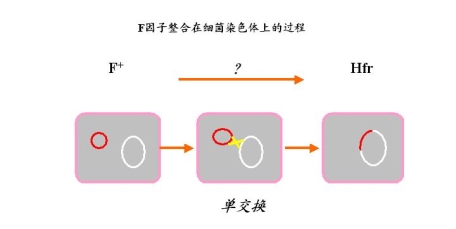
F因子——又称F质粒(是质粒中的一种)
质粒——染色体外遗传因素可自主复制
有的可以整合也可独立——附加体(episome)
为什么可以整合到“染色体”上?与其特殊的结构有关
为何可以插入不同的位置?
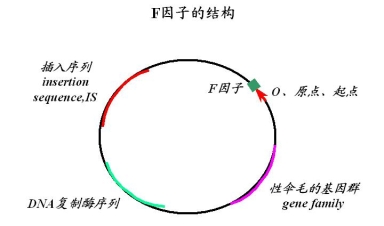
F因子的结构包括以下几个部分:
1)原点——转移的起点
2)形成性伞毛的基因群(gene familu)
——使细菌表面出现一至若干个性伞毛,通过其与F-细菌接合
3)DNA复制酶基因——与F因子本身的复制有关
4)插入序列(insertion seguence IS)
与其插入细菌染色体的过程有关
①F因子中IS与细菌“染色体”的IS配对发生交换
②IS有极性向左、向右
六、细菌的交换过程
原核生物——细菌
来自供体的部分基因组(外基因子)
来自F-的完整基因组(内基因子)
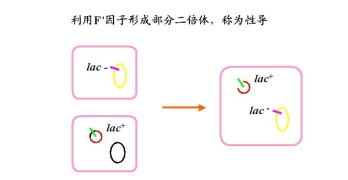
部分合子(merozygote)又称部分二倍体(partial diploid)交换的可能性一次交换或奇数次交换
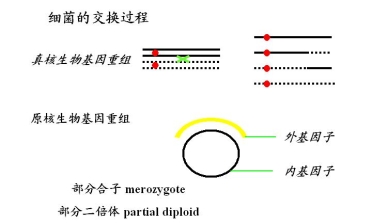
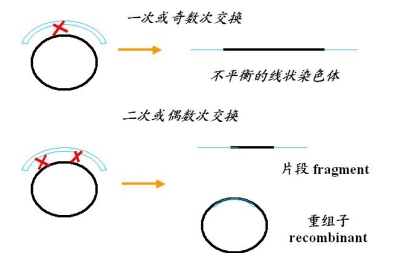
细菌重组的特点:
① 偶数交换才能产生平衡的重组子
② 相反的重组子(reciprocal recombinant)不出现,所以选择培养基上只出现一种重组子
七、重组作图——重组作图法(recombination mapping)
中断杂交试验中,基因转移的先后次序,以时间为单位——基因连锁关系
取样时间不可能近到以秒来计,所以距离太近的两基因间定位用中断杂交试验技术——不可靠
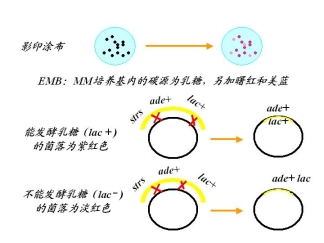
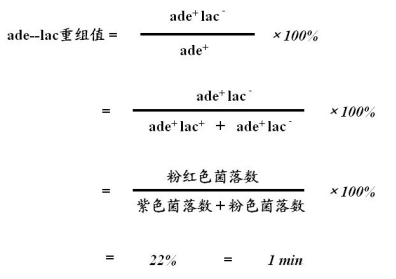
1个时间单位(1分钟)≈20%重组值
八、性导sexduction
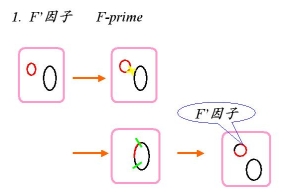
F’因子特点带有F因子
2) 带有核“染色体”的部分片段
3) 独立存在,也可以在原位重新整合,形成Hfr菌株
4) F’×F—→ F’ + F’ ,但发生重组的频率比Hfr×F—低
对于基因重组的频率而言是非常有意义的
2.性导
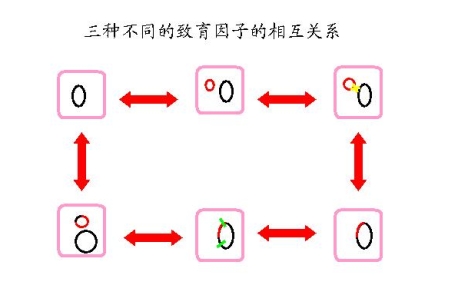
3.三种不同的致育因子的相互关系
第三节噬菌体的遗传分析
一、关于噬菌体baiteriophage简作phage
1. 噬菌体
 是一类在活细菌内生活的病菌或能够侵梁细菌的病菌,无细胞结构,由核和蛋白质外壳构成
是一类在活细菌内生活的病菌或能够侵梁细菌的病菌,无细胞结构,由核和蛋白质外壳构成
2.裂性噬菌体virulentphage使主细菌发生裂解的phage
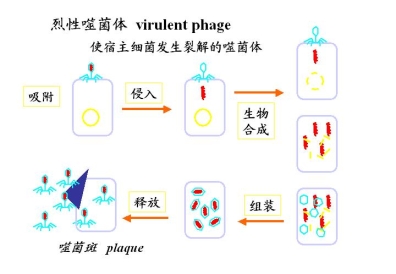
噬菌斑(plague)
plague侵染细菌使细菌的菌落(不透明菌)出现一个圆形的透明区,即plague
噬菌斑,依基因型不同有大、有小
3.溶源性lysogeny
几个概念:溶源性温和噬菌体temperate phage
溶源性细菌或称溶源菌lysogenic bacteria
前噬菌体或原噬菌体prophage
两种原噬菌体的存在方式
溶源性:有些细菌带有噬菌体(某种)但并不立即导致溶菌,这种现象称为溶源性
溶源性细菌:带有某些噬菌体的细菌,不出现溶菌现象称为溶源性细菌
温和噬菌体:侵染细菌不引起溶菌现象的噬菌体称为温和噬菌体
原噬菌体:细菌细胞内含有的无感染能力的噬菌体
溶源性细菌的特征:①免疫性 ②外异偶然因素(如紫外线、雷素C)诱导释放噬菌体
二、噬菌体的基因重组
![]()
![]() 野生型phage → 突变型
野生型phage → 突变型
→ 突变型
举例:裂性噬菌体T2的两对性状及其重组
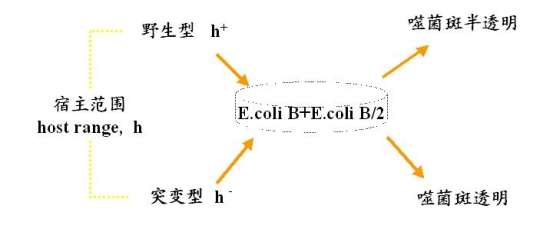
两对性状之一:有关缩主范围(hostrange,h)
两对性状之二:有关噬菌斑形态
① 缩主范围:
野生型h+ 突变型h-
可以侵染和裂解E.coli.B 可以侵染和裂所E.coli.B
不可以浸染E.coli.B/2 可以侵染和裂所E.coli.B/2
(因为表面有阻止吸附的结构) (缩主范围扩大了)
②噬菌斑形态plague morphology
野生型r+ 快速溶菌的突变型r-
小噬菌斑ф 1mm左右 大噬菌斑ф 2mm左右
(溶菌阻碍现象引起)
实验过程
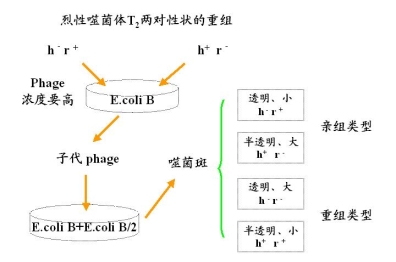
对于不同的快速溶菌类型有三种ra rb rc
三组杂交 1)rah+ rah-
2)rbh+ r+h-
3)rch+ r+h-
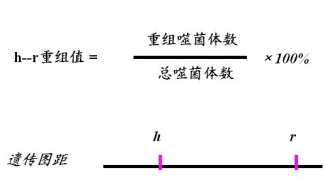
计算重组值,画连锁图
三、转导transduction
以噬菌体为媒介将细菌的小片段染色体或基因从一个细菌转移到另一个细菌的过程
1.普遍性转导generalized transduction
这是和种通过完全缺陷噬菌体(或称假噬菌体)将供体菌中的任何基因(包括质粒)携带进入受体菌细胞中,并与受体菌“染色体”同源区段配对,经双交换整合在“染色体”上,使受体菌实现各种性状的转导
Ⅰ.转导实验:
鼠伤寒沙门氏杆菌styphimurium
供体菌 野生型如LT22A(trp+)
受体菌 营养缺陷型LT22A(trp-)
转导媒介P22噬菌体——烈性噬菌体
1.1 P22噬菌体浸染供体菌,释放烈解物的过程
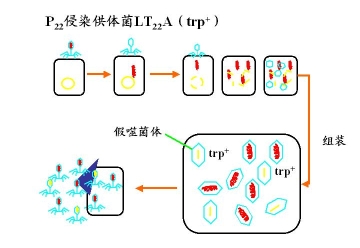
1.2 裂解物再感染受体菌,发生基因重组的过程
少量裂解物+大量受体菌 每个受体菌表面最多只有一个phage侵染
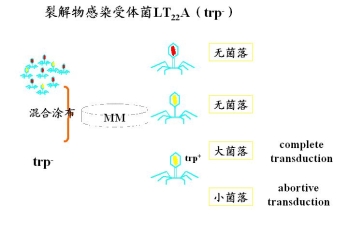
根据计算非选择性标记的频率,从而确立几个基因的相对位置及片段的长度
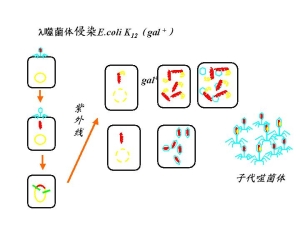
2.特异性转导specialized transduction
又称局限性转导restricted transduction媒介载体入噬菌体(一种温和噬菌体)
供体E.col:k12.gal+
受体E.col:k12.gal-