第九章 遗传物质的改变(一)
染色体畸变
计划学时 8
![]() 染色体畸变 chromosomal variations/aberrations
染色体畸变 chromosomal variations/aberrations
染色体结构变异
遗传物质的改变 染色体数目变异
基因突变 genetic mutations 又称点突变point mutations
第一节 染色体结构的改变
起因 染色体或染色单体的断裂(breakage)
- 无着丝粒的片断(segment)最后丢失
- 原位重新结合或重建(restitution)
- 非重建愈合(nonrestitution union)
所以有:
- 缺失 decetion或deficiency
- 重复 duplication或 repeat
- 倒位 inversion
- 易位 translocation
一. 研究染色体畸变的几种好材料
- 玉米2n=20,10对同源染色体,
10条染色体形态上可以相互区别
RD:粗线期 同源染色体联合配对,细长的染色丝
- 果蝇 2n=8
唾腺染色体又称多线染色体polytene chromosome 又称巨大染色体
1) 同源染色体的体细胞联合
2) 细胞DNA复制,但着丝粒不分裂,无细胞数目增加,有染色体周期
3) 横纹相对大小,空间排列是恒定的
4) 唾腺内
二. 缺失 deficiency 指染色体的某一区段丢失
1. 缺失的类别
顶端缺失 terminal deficiency ——丢失的染色体区段为某臂的外端
中间缺失 interstitial deficiency——染色体缺失的区段可能是某臂的内段
断片 fragment 缺失的区段,无着丝粒,称为断片或片段segment
缺失杂合体deficiency heterozygote 某个体的体细胞内含有正常的染色体及其缺失染色体,称为缺失杂合体
缺失纯合体 deficiency homozygote 某个体的体细胞内的缺失染色体是成对的,称为缺失纯合体
- 缺失的细胞学鉴定
同源染色体配对时出现缺失环 deletion loop
如:黑腹果蝇幼虫唾腺染色体
3. 缺失的遗传学效应
(1)缺失纯合体难以存活
(2)缺失杂合体
![]() 正常染色体 雌雄配子均可育
正常染色体 雌雄配子均可育
配子
缺失染色体 雌配子可育,但生活力弱,雄配子不育
所以,缺失杂合体雄花粉半不育,胚囊可育
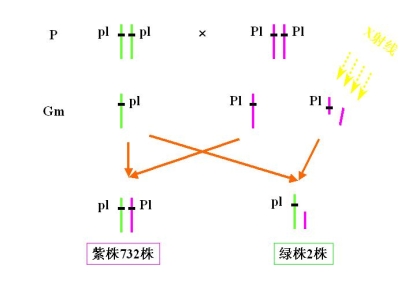
(3)假显性现象 pseudodominance
例:玉米植株颜色有关的一对基因Pl(紫茎)和绿茎pl缺失遗传效应
Pl、pl位于第六号染色体长臂外端
例:人类的猫叫综合症“Cridu-chat” syndrome
第五号染色体短臂缺失(5P-)
两眼距离较远,耳位低下,智力迟钝,生活力差
明显特征:哭声轻,音调高,似猫叫
三. 重复 duplication
是指染色体多了自己的某一区段
1. 重复的类别
顺接重复 tandem duplication 是指染色体上重复区段与原来正常染色体的顺序相同
反接重复 recerse duplication 是指染色体上重复区段与原来正常染色体的顺序180度倒转
重复纯合体 duplication homozygote
重复杂合体 duplication heterozygote
2. 重复的细胞学鉴定
同源染色体联合对产生重复环 duplication loop
3. 重复的遗传学效应
主要影响是:扰乱了基因的固有平衡体系
(1)剂量效应 dosage effect
(2)位置效应 position effect
果蝇X染色体 16区A段 红眼――椭球形,
——小眼数目780个左右
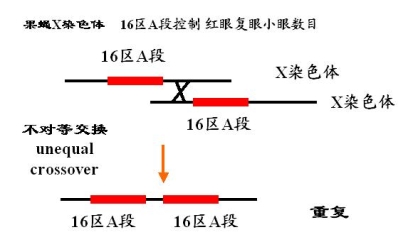
正常 b/b 正常 780个小眼
杂合棒眼Bar B/b 棒眼 358个小眼
纯合棒眼 B/B 棒眼 69个小眼
杂合重棒眼 Bb/b 重棒眼 45个小眼
纯合重棒眼 Bb/Bb
剂量效应 由于重复区段数目的增加而产生的效应影响
位置效应 重复区段排列方式的不同而引起的遗传效应差异
四. 倒位 inversion
是指染色体的某一区段断裂,发生180度倒转并重新连接
(一) 倒位的类别
臂内倒位 paracentric inversion 发生倒位的区段不包括着丝粒,在一条臂内发生
臂间倒位 pericentric inversion 包括着丝粒在内的带有两条臂的一个区段发生的倒位
倒位纯合体
倒位杂合体
(二)倒位的细胞学鉴定
倒位区段的长短决定 
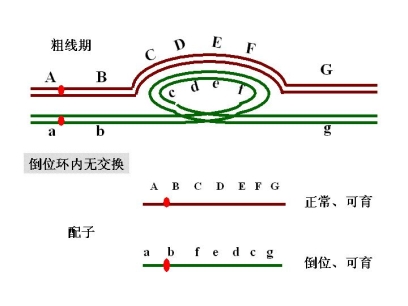
倒位环 inversion loop
(三) 倒位的遗传学效应
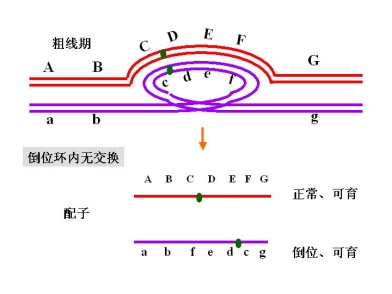
1. 倒位杂合体产生部分败育配子
倒位(臂内)杂合体
如果不发生交换,产生两种配子
如果倒位环内发生一次单交换
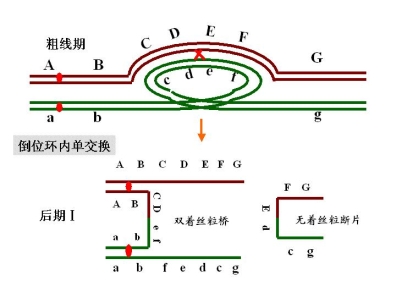
产生配子:a b c e d f 正常,可育
![]()
![]() a b c
a b c
a b d e
a b e d c f g 倒位,可育
如果倒位环内发生一次双交换
产生配子:a b c d e f g 正常可育
a b c d e f g 重组正常可育
a b e d c f g 重组倒位可育
a b e d c f g 倒位可育
臂间倒位杂合体
如果无交换发生 产生配子
a b c d e f g h 正常可育
a b f e d c g h 倒位可育
如果倒位环内发生一次单交换,产生配子
a b c d e f g h 正常可育
![]()
![]() a b c d e f b a
a b c d e f b a
h g f e d c g h
a b f e d c g h 倒位可育
如果倒位环内发生一次双交换,产生配子
a b c d e f g h 正常可育
![]()
![]() a b c d e f g h
a b c d e f g h
a b f e d c g h
a b f e d c g h 倒位可育
所以,
1. 倒位杂合体产生部分败育配子 部分不育 partial sterility
2. 倒位杂合体的重组值大大降低 只有双交换才能产生重组类型配子
3. 物种进化的因素
五. 易位 translocation
是指某染色体的一个区段转接在非同源的另一染色体上。
(一)易位的类别
单向易位 simple translocation 单方向转移片断
相互易位 reciprocal translocation
易位纯合体
易位杂合体
(二)易位的细胞学鉴定
单向易位杂合体 RD偶线期 “T”
相互易位杂合体 RD偶线期 “+”
(三)易位的遗传学效应
相互易位杂合体 联合
![]()

后期I 同源染色体随机分离
后期I 1/2adjacent segregation 邻近分离
配子 缺失 重复 不育
1/2alternate segregation 交互式分离
![]() 正常 可育
正常 可育
配子
易位 可育
1. 易位的杂合体的配子半不育性
2. 易位的杂合体自交后代的表现类似于带有一对杂合基因个体自交后代的表现
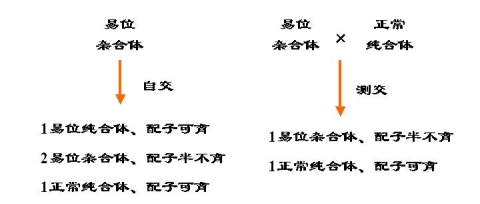
3. 易位可改变原来的连锁群
4. 物种进化的因素
六. 染色体结构变异的应用
(一)利用缺失进行基因定位
关键:
1)使载有显性基因的染色体发生缺失,让它的隐性等位基因有可能表现“假显性现象”
2)对表现假显性现象的个体进行细胞学鉴定,发现某染色体缺失了一段(某一区段),这就说明,该显性基因及其等位的隐性基因位于该染色体的缺失区段。
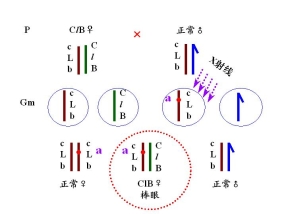
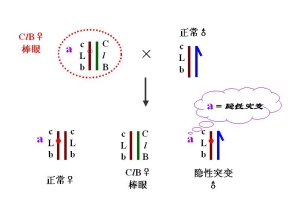
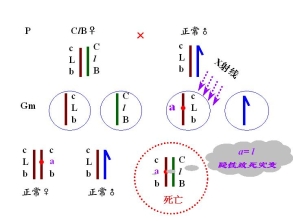
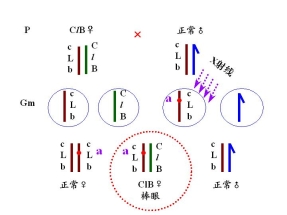
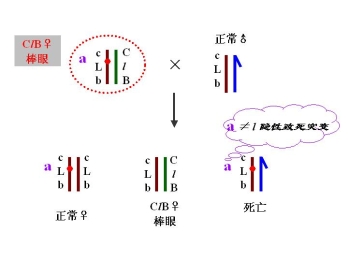
(二)果蝇的CLB测定法
――鉴别果蝇X染色体的隐性突变
CLB品系:平衡致死品系 balanced lethal system
永久杂质 permanent hybrid
雌性的杂合体
![]()
![]()


C 位于X染色体上的打倒位 倒位降低交换值
——倒位杂合体 配子中无重组配子
l 位于X染色体上的隐性致死基因
——具有ClB:雄性不成活
B 位于X染色体上的显性棒眼基因
——16区段内的重复 表型上的鉴别标记

有一雄性果蝇发生隐性突变
鉴定1. 隐性突变是否发生在X染色体上
2. 隐性突变是否是致死突变 与l等位、与l非等位
3. 突变率
鉴定:
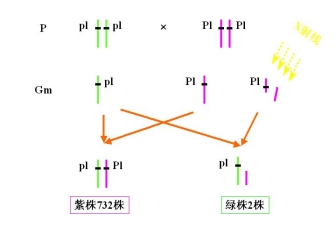
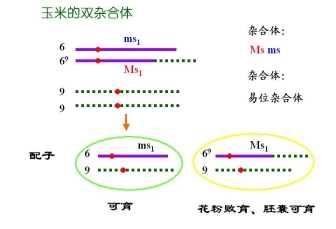
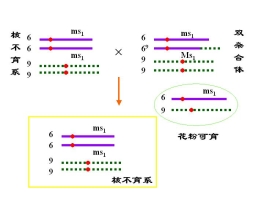
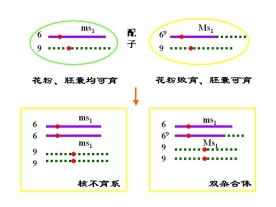
(三)利用易位创造玉米核不育的双杂合体保持系
等位基因 MS 雄性可育基因――雄性可育
ms 雄性不育基因――雄性不育
P msms × MSMS
![]() 雄不育 雄可育
雄不育 雄可育
F1 MSms雄性可育
F1的雄性可育,实际上是雄性不育株的不育性不能得到保持
为此,利用易位创造玉米核不育系的双杂合体保持系
玉米的双杂合体

第二节染色体数目的变异
一、染色体组及其倍数的变异
1.染色体组genome
染色体组genome
把基数的X个染色体总一起来称为一个组即染色体组
配子中含有的染色体数称染色体组
2n=体细胞中chr数目
n=性细胞中chr数目
基数:一个染色体组内含有的染色体数,用X表示
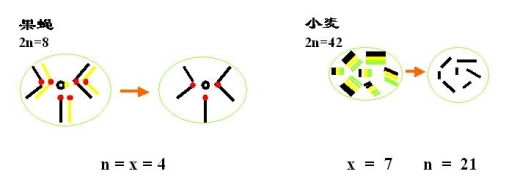
二倍体果蝇2n=n8 n=4 2n=2x=8 n=x=4
多倍体小麦2n=4x 2n=6x=42 n=3x=21 x=2
染色体组的基本特征:
同一个染色体组的各个染色体的形态,结构和连锁基因群都彼此不同,但它们却构成了一个完整的协调体系,缺少其中任何一个都会造成不育或性状变异。
2.染色体数目变异的种类
(1)整倍体euploid体细胞内含有完整的染色体组的类型
二倍体diploid 2n=2x
一倍体monoploid 2n=x
单倍体haploid 2n=n
![]()

一倍体与单倍体的区别:
多倍体polyploid 2n=3x 4x 5x 6x
同源多倍体autopoluploid
多倍体中,如果加倍的染色体组是来自同一物种,或在原有染色体组的基础上加倍而成的,称为同源多倍体2n=AAAA 2n=AAA 2n=AAAAAA
异源多倍体allopolyploid多倍体中,如果增加的染色体组来自不同的种属称为异源多倍体
(2)非整倍体aneuploi
d在二倍体的基础上,增加或减少一个或多个chr的类型
超倍体hyperploid 2n+
三体trisomic2n+1 四体tertrasomic2n+2 双三体doubletrisomic2n+1+1
亚倍体hypoploid2n-单体monosomics2n-1 缺体nullisomic2n-2 双单体doublemonosomics2n-1-1
二、 整倍体euploid
1.单倍体haploid
▲一些低等植物的配子体是单倍体——正常
一些昆虫的雄体(如蜂类等膜翅目昆虫)是单倍体——正常
▲高等植物单倍体与二倍体相比形态特征体形弱小,(包括根、茎、叶、花等器官都很小)有时出现黄化苗
▲单倍体的高度不育
原因:无同源染色体联合配对,所以染色体无规律地分离到不同中去。
▲单倍体的利用
利用F1花粉培养成单倍体植株又以获得广泛变导师的个体,隐性突变可以表达
单倍体植株人工加倍,可获得育性正常的纯分个体缩短育种年限
人工诱变单倍体,提高诱变效果
2.同源多倍体autopolyploid
香蕉是同源三倍体
马铃薯是同源四倍体
甘蔗是同源六倍体
▲同源四倍体与二倍体比较有以下特点
①一般表现为形态的巨大性细胞粒增大细胞体积增大叶片上气孔增大,花粉粒增大,花果、果实增大细胞数目未见增多
②生长发育缓慢,开化,成熟较迟,适应性较强
③同源多倍体一般都表现各种程度的不育
所有的染色体均为2/2分离则n=2x可育
或所有的染色体均发出3/1分离则n=3x n=x可育
或有的3/1有的2/2则n=???不育
如:四倍体蔓陀罗Daturastramonirm4n=48n=2x=24可育
四倍体番茄4n=48n=2x=24可育
④同源四倍体的遗传表现比二倍体复杂
一对等位基因A a
二倍体中基因型有三种AA Aa aa
同源四倍体中基因型有五AAAA种四式 AAAa三式 AAaa二式 Aaaa单式 aaaa零式
同源四倍体的各种杂合体数及后代表型比数
同源三倍体
![]() 来源 四倍体 二倍体
来源 四倍体 二倍体
同源三倍体
同源三倍体育性很低 联合 后期Ⅰ分离
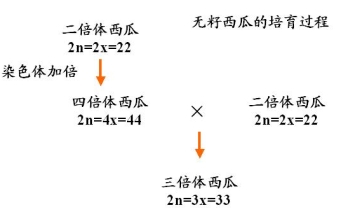
例:香蕉、黄花菜、水仙、无籽西瓜
3.异源多倍体
萝卜甘蓝的属间杂种多倍体——异源四倍体
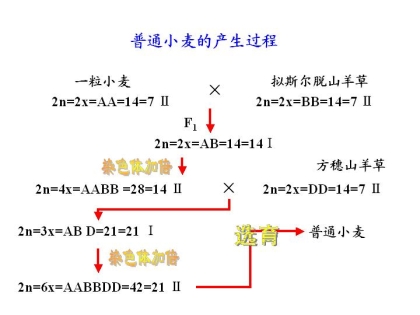
说明:一粒小麦 → 二粒小麦 → 普通小麦的溶化过程
异源多倍体的产出是种间杂种染色体加倍的结果
性状遗传规律同二倍体
4.多倍体的诱发
高温、低温、离心、超声波、嫁接
秋水因素:淡黄色粉末、针状结晶、易溶于凉水、有剧毒,来自秋水仙的鳞芳和种子
分子式:C22 H25 O6 N + 1.5 H2O
阻止纺锤丝形成
0.01%--1.0%
作用:抑制细胞分裂时锤体的形成
5.多倍体的应用
四倍体番茄=2倍的二倍体番茄
三倍体甜菜,含糖量高,耐寒,产量高
三倍体杜鹃花、花期长、花大
无籽西瓜
八倍体小黑麦:
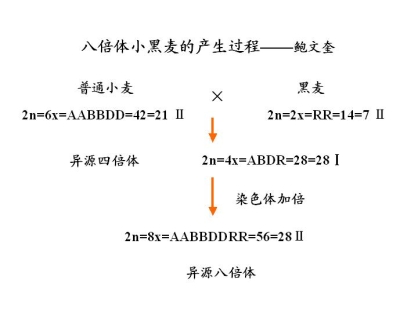
八倍体小黑麦:
产量高、抗逆性和抗病性强
耐瘠耐寒、面粉白、蛋白质含量高
发酵性能好、茎杆可作青饲料
三、非整倍体
1.单体和缺体
▲单体和缺体对生物体的影响较大
在二倍体(2n=2x)植物中单体和缺体大多不能成活
原因:是整个染色体组的基因平衡问题
▲在多倍体中,特别是异源多倍体中,单、缺体较为常见,
原因:多倍体植物的一个染色体组内缺少个别染色体引起的基因不平衡可以由其他染色体组的完整而得到补偿。
单体产出配子
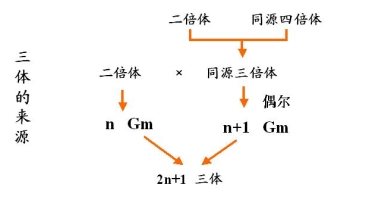
2.三体
三体在二倍体植物中常见
三体来源
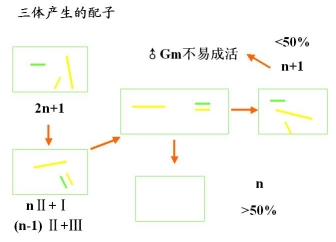
三体的基因分离
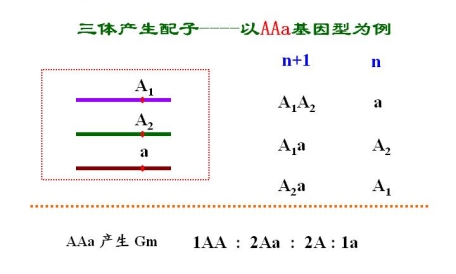
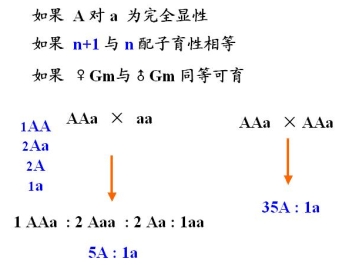

3.人类的非整倍体
先天愚型
病症:头颅前后经短、枕骨扁平、眼小、两眼外侧高而内侧低、鼻梁扁平且宽、口半张、舌常伸出口外,舌有龟裂通贯手、发育迟缓、智力低下,平均寿命短
Patau综合症
Edwards综合症
病症:头小而长、大囟门,鼻梁窄而长、小嘴腭狭窄、耳低位、短颈、平均寿命6个月
发生原因:配子形成过程中染色体不分开
4.非整倍体的应用:
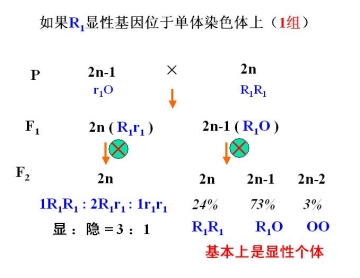
①利用单体进行基因定位,确定基因所在的染色体
例:如何用单体确定小麦红粒基因位于哪一条染色体上?
小麦 2n = 6x = 42——适于用单体进行分析
小麦单体系统有 21 个

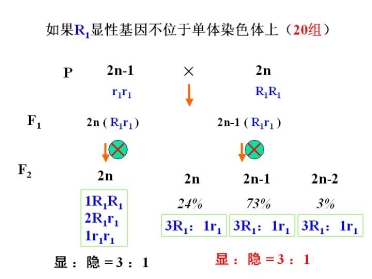
②利用二体进行基因定位——确定基因所在的染色体
例:水稻上发现了一个矮杆突变体,表现为单基因隐性遗传,如何利用水稻的非整倍体把它定位到相应的染色体上?
已知:水稻 2n = 2x = 24 适于用三体进行分析
水稻三体系统有 12 个
原理:三体为母本或父本时,产出的配子类型和比例不同

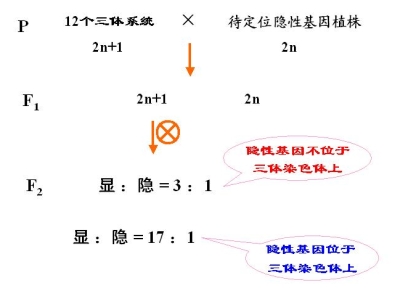
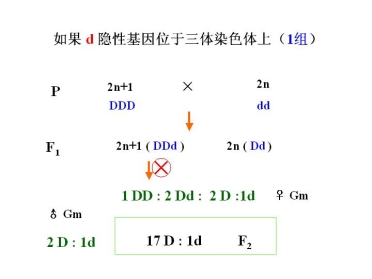

相似的问题:
A:水稻隐性基因能否用三体的测交法确定?
B:小麦的隐性基因突变如何用单体进行基因定位?
C:如何将水稻的显性基因定位于特定的chr上?
D:图解说明如何用单端二体把小麦的红皮基因定位到相应的染色体上(或定位到特定的臂上)
③利用三体配制大麦一代杂种
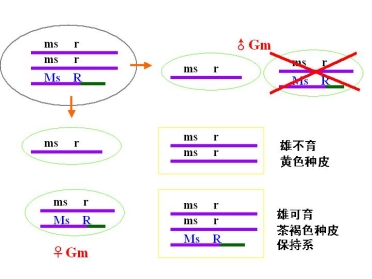
作业:思考题